15 - Mouth, Nose, and Paranasal Sinuses
Editors: Mills, Stacey E.
Title: Histology for Pathologists, 3rd Edition
Copyright 2007 Lippincott Williams & Wilkins
> Table of Contents > VI - Thorax and Serous Membranes > 20 - Normal Heart
20
Normal Heart
Gerald J. Berry
Margaret E. Billingham
Introduction
With the introduction of the cardiopulmonary bypass technique into clinical practice a half-century ago, pathologists found themselves dealing more frequently with cardiac specimens in the surgical pathology laboratory. For many years this consisted of explanted valves, pericardiectomy specimens, ventricular aneurysectomy excisions, and interventricular myomectomy specimens. It was not until cardiac transplantation began in 1968 that the surgical pathologist began handling entire heart specimens. Currently, more than 73,000 heart and combined heart-lung transplant procedures have been recorded in the registry of the International Society of Heart and Lung Transplantation (1). Thoracic transplantation in the pediatric age group ranging from neonates to adolescents is now commonly performed (2). The explanted hearts from infants and children often have complicated congenital heart lesions that have undergone one or more corrective surgical interventions. Surgical pathologists need to have some expertise in the
P.528
morphologic evaluation of end-stage heart disease of all types. Since the early 1970s, endomyocardial biopsies for the assessment of allograft rejection, infiltrative lesions, and inflammatory disorders are performed routinely. Today many institutions have initiated ventricular assist device (VAD) programs, and ventricular apical specimens are routinely submitted. Like other biopsy specimens, these fall under the auspices of the surgical pathologist. Further, the introduction of complex electrophysiologic mapping techniques and imaging modalities such as transesophageal echocardiography, intravascular ultrasound, cardiac gated spiral-computed tomography (CT) angiography, and magnetic resonance imaging (MRI) have renewed interest in structural-functional correlations. Pathologists have assumed a central role in the clinical evaluation and application of these technologies.
The purpose of this chapter is to review normal cardiac anatomy and histology. The histology of the great vessels is described elsewhere in this book. For the most part, adult histology is described; but, where important differences exist, the histology in infants and children is also addressed. Furthermore, the purpose is not to be encyclopedic. Rather, we will highlight those areas that are of practical importance to the surgical pathologist to enable the distinction between normal and subtle cardiac pathology. As each of the major anatomic divisions of the heart is described, relevant aspects of aging, gender changes, and/or applied anatomy are mentioned. We have also included recommendations for handling endomyocardial biopsy specimens and common artifactual alterations in this discussion.
Heart Weights
The weight of the normal adult heart is generally achieved by the end of the second decade of life. Heart weight in infants and children is related to age and body size, and tables are readily available in published works (3,4). A variety of approaches to estimating the weight of the adult heart have been used in clinical practice. These differ in complexity and accuracy; for example, the practical guidelines for unfixed adult heart weights published by Hudson in 1965 (5) indicate the male heart weighs 0.45% of the body weight (325 75 g; average 300 g) and the female heart weighs 0.40% of the body weight (275 75g; average 250 g). Heart weight varies with age, sex, and body height and weight. Using this type of approach, the heart weight in young athletic adults may approach or exceed the upper limits of normal by up to 25%, whereas in the elderly it may approach or be slightly below the lower limits of normal.
Kitzman and colleagues from the Mayo Clinic (6) proposed a more comprehensive approach that predicts the normal heart weight of a formalin-fixed specimen based on body weight. In their study, body height as a predictor proved less accurate than body weight. The impact of formalin fixation on the weight of the heart has been addressed with conflicting findings. In one study, the overall finding was that heart weight increased by 5% after fixation, although considerable individual variation ranging from weight loss to weight gain was reported (7). Descriptive tables of normal ventricular wall thickness, chamber size, and valvular orifice measurements are available in textbooks and postmortem dissection manuals (4,8,9,10).
Prenatal Fetal Circulation
It is beyond the scope of this chapter to discuss cardiac embryology. Developmental molecular biology has provided many new insights in cardiogenesis, some of which have radically altered previously held tenets (11,12,13,14). Currently it is thought that the human heart begins to beat around day 17. Well-oxygenated blood leaves the placenta via the umbilical vein, a portion of which passes through the hepatic sinusoids, and the remainder bypasses the liver by way of the ductus venosus to enter the inferior vena cava (IVC). Caval blood enters the right atrium together with an admixture of deoxygenated blood from the lower body. Approximately one-third of this blood is diverted through the interatrial septum (ostium secundum) into the left atrium, where it mixes with the small amount of deoxygenated blood returning from the lungs. The blood then passes into the left ventricle and exits into the ascending aorta to supply the coronary arteries, brain, and upper limbs. Approximately 50% of the blood is returned to the placenta for reoxygenation via the umbilical arteries. The remainder of the blood supplies the lower half of the body. Blood returning to the heart from the upper body reaches the right atrium through the superior vena cava (SCV). It mixes with the residual two-thirds of the blood from the IVC and enters the right ventricle. A small portion is distributed to the lungs, but the majority is diverted across the ductus arteriosus to the descending aorta. Blood is returned to the placenta with only a small amount reaching the lower body of the fetus.
Postnatal Circulation
At birth, a series of anatomic and physiologic changes occur, beginning with the infant's first respiratory efforts. The circulation of fetal blood through the placenta ceases, and the infant's lungs expand. As alveoli become aerated, the pulmonary vascular bed dilates, causing increases in pulmonary blood flow and abrupt decline in pulmonary vascular resistance. Additionally, active remodeling of the pulmonary vasculature begins and continues over a period
P.529
of a few weeks. This consists primarily in the reduction of the muscular medial layer of arterioles and small arteries. Increased blood return to the left atrium by way of the pulmonary veins results in slightly higher left atrial pressures compared to the right atrium. This causes the flap covering the foramen ovale to seal, leaving the indentation called the fossa ovale. By the end of the first month after birth, the significant differences in ventricular hemodynamic load result in thickening of the left ventricular wall and thinning of the right ventricular wall. The ductus arteriosus, located between the left pulmonary artery and aortic arch, functionally closes within 15 hours after birth, and the umbilical veins begin to constrict.
Pericardium
The pericardium surrounds the heart and consists of a fibrous and a serous sac. The fibrous, or parietal, pericardium envelops the heart and is reflected off the ascending aorta, the pulmonary trunk, and the terminal 2 to 4 cm of the SVC, the distal segment of the IVC, and the pulmonary veins. In the normal state, the fibrous pericardium surrounding the heart remains unattached to the serous (visceral) pericardium except at the pericardial reflections. The parietal pericardium is composed of collagenous fibrous tissue (Figure 20.1). Its inelastic tendencies account for the development of cardiac tamponade when it is acutely stretched with more than 250 ml of fluid. It may contain variable amounts of adipose tissue, particularly toward the apex of the heart. A thin layer of mesothelial cells on its inner surface lines the parietal pericardium. It is normally less than 1 mm in thickness. Heterotopic tissues within this layer include thyroid and thymic elements. Neoplastic and hyperplastic alterations have been described, including intracardiac thyroid in the right ventricular outflow tract (15).
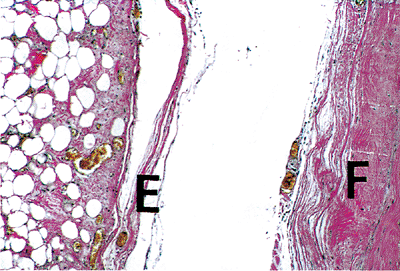 |
Figure 20.1 Section showing fibrous pericardium (F) separated from the thinner epicardium (E) (elastic van Giesen). |
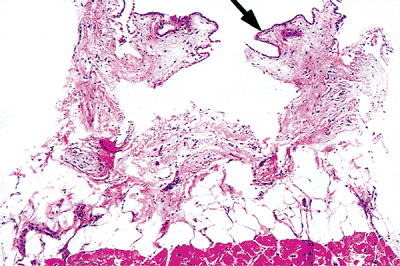 |
Figure 20.2 Section showing the epicardium lined by mesothelial cells (arrow) and covering the subepicardial adipose tissue and myocardium (H&E). |
The serous, or visceral, pericardium is also called the epicardium of the heart. It is a single layer of mesothelium that envelops the heart and is in continuity with the fibrous pericardium at the pericardial reflections at the great vessels (Figure 20.2). This delicate membrane covering the heart contains variable amounts of adipose tissue within which are embedded the coronary arteries and veins, lymphatic vessels, nerves, fibroblasts, and macrophages (Figure 20.3). The normal epicardium often has small aggregates of lymphocytes that are present from birth.
Between the two mesothelial layers of the parietal and visceral pericardium is a potential space containing up to 50 mL of straw-colored fluid that allows the surfaces to glide over one another in the normal state. The histologic composition of the two layers of the pericardium does not alter with age and is similar in infants, children, and adults.
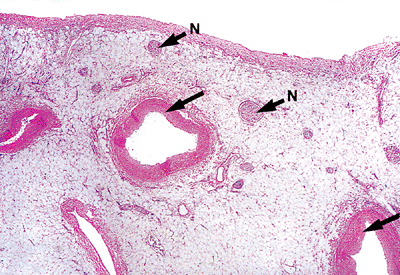 |
Figure 20.3 Section of the epicardium showing the relationship of coronary vessels (arrows) and nerves (N) to the epicardium (H&E). |
P.530
Applied Anatomy
Surgical pathologists may receive pericardiectomy specimens in cases of constrictive pericarditis or recurrent tamponade. Nonspecific fibrosis and chronic inflammation are observed, but granulomatous infection and neoplastic infiltrates should be excluded. In explanted hearts, the pericardial surfaces are often thickened by previous disease states or surgical interventions, in which case the epicardium and parietal pericardium are adherent. Occasionally nodular collections of epicardial fat may be misinterpreted grossly as metastases. Congenital cysts of the pericardium are rare. Heterotopic thyroid and thymic tissue should be distinguished from cardiac involvement by contiguous spread or metastasis. In obese subjects, there may be excessive epicardial deposits of adipose tissue, and the measurement of ventricular thickness should be carefully evaluated. In conditions of cachexia, epicardial fat may undergo serous atrophy, resulting in small gelatinous tissue tags (10).
Cardiac Skeleton
The cardiac skeleton at the base of the heart is the central supporting structure to which most of the fibers of the myocardium are attached and to which the atrioventricular and aortic valves are anchored. A separate mass of fibrous tissue (the conus ligament) joins the pulmonary ring to the aortic ring. It also serves to separate the atrial and ventricular chambers. It is composed of the right and left fibrous trigones, membranous septum of the interventricular septum, and the fibrous annuli of the atrioventricular foramina (16). The fibrous skeleton is composed of layers of dense collagenous connective tissue admixed with small numbers of elastic fibers and, on occasion, small aggregates of adipose tissue. The fibrous tissue of the mitral and aortic valve rings is more substantial than the right-sided valves, and the right and left trigone create the direct mitral-aortic continuity (17). The right trigone and membranous septum form the central fibrous body within which the atrioventricular bundle is embedded. Bone or cartilage is not normally found in the fibrous skeleton of human hearts, although they have been described in some animal species.
Applied Anatomy
Portions of the cardiac skeleton may be seen in explanted valves. The close proximity of the fibrous skeleton to the conduction system and valvular leaflets is an important consideration for surgeons. Alterations can significantly affect the functional outcomes of all these components.
Internal Structure of the Heart Wall
The wall of the heart in all chambers consists of three main layers: (a) the endocardium, (b) the intermediate muscular portion of the myocardium, and (c) the external portion, or epicardium. Most investigators believe that the endocardium is homologous with the tunica intima of blood vessels, the myocardium with the smooth muscle media, and the external epicardium with the adventitial layer. The endocardium consists of a single layer of endothelial cells (Figures 20.4,20.5) with a subendothelial portion containing a loose elastic framework and collagen bundles, as well as nerves and delicate blood vessels (Figure 20.6).
While the chambers of the heart are composed of these three layers, there can be variation in the thickness of one or more components. A rudimentary layer of smooth muscle is often found in the endocardium of both atria and the
P.531
ventricles (Figure 20.7). The endocardium of the atria is thicker than the ventricles. Small bundles of smooth muscle are often found but are of questionable functional significance. The myocardium is composed of bundles of myocytes separated by fibrous bands (Figure 20.8). The individual myocytes form a syncytium with end-to-end junctions (called intercalated disks) and sometimes side-to-side junctions. Individual myocytes contain a central ovoid nucleus with a clear zone at the poles (Figure 20.9). The normal cardiac myocyte contains small amounts of lipofuscin granules (lysosomes), which increase in quantity with age. The myocytes are filled with contractile myofibrils (actin and myosin). In addition, atrial myocytes contain atrial dense-core bodies that are demonstrable by electron microscopy (Figure 20.10). The epicardial portion of the chambers was described earlier.
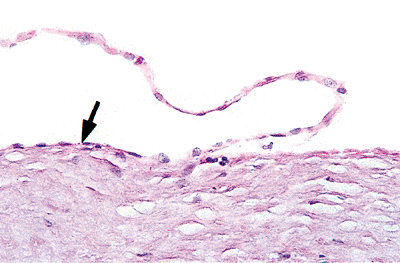 |
Figure 20.4 Section showing a single layer of endothelial cells covering the myocardium (arrow) (H&E). |
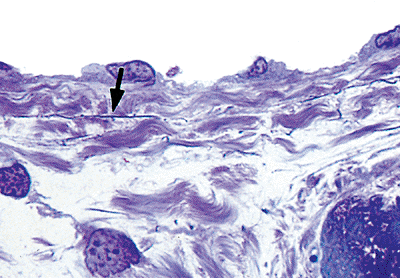 |
Figure 20.5 One- m thick section (plastic embedded) of the endothelium covering the fenestrated elastic fibers (arrow) and collagen fibers of the subendocardium (toluidine blue). |
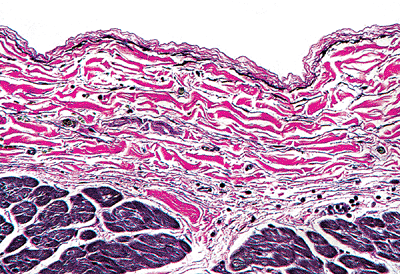 |
Figure 20.6 Section of the endocardium showing the distribution of the elastic fibers (black) and the collagen bundles (red), as well as vessels and nerves in the subendocardial layer. |
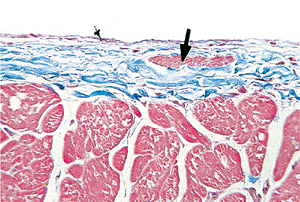 |
Figure 20.7 Section of endocardium showing smooth muscle bundle (arrow) (Masson's trichrome). |
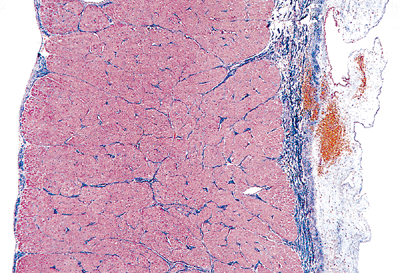 |
Figure 20.8 Section through the right ventricular wall showing the distribution of collagen from the endocardium (left) through the myocardium and to the epicardium (right) in the normal heart (Masson's trichrome). |
Interatrial Septum
The interatrial septum separates the right and left atrial chambers. In the normal state this separation is complete, although the foramen ovale may retain a small communication at its floor where the septum has not fused (the so-called patent foramen ovale). This is reported in up to 30% of adults and is usually without clinical complications (18). The foramen ovale is closed by the septum to form the circular, dime-sized oval fossa. This represents the true interatrial septum, with the remainder of the anatomic structure being comprised of caval or pulmonary venous infoldings and the muscular and membranous components of the atrioventricular region (17). The membrane covering the oval fossa is a paper-thin, translucent layer early in life that becomes fibrotic and thickened with age, measuring up to
P.532
2 mm in thickness. The histology of the septum displays variable amounts of atrial muscle, fibrous tissue, mature adipose tissue, and, on occasion, brown fat (Figure 20.11). The thickness of the atrial septum varies considerably.
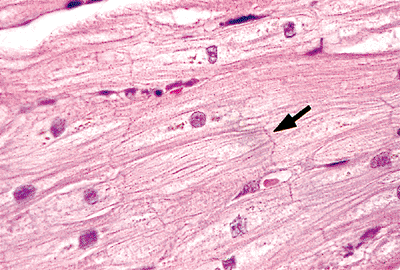 |
Figure 20.9 Section of myocardium showing ovoid nuclei and intercalated disks (arrow) (H&E). |
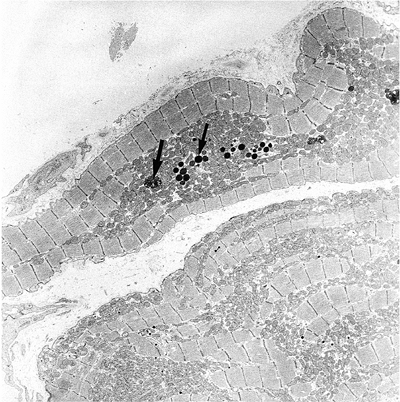 |
Figure 20.10 Electron micrograph of atrial dense-core granules (small arrow) that can be differentiated from the lipofuscin granules (large arrow). |
Applied Anatomy
Lipomatous hypertrophy of the interatrial septum is characterized by accumulations of adipose tissue within the U-shaped lip, or limbus, of the oval fossa and may produce bulging of the interatrial septum. In some patients, the patent foramen ovale can be the site of interatrial shunting or paradoxical embolism.
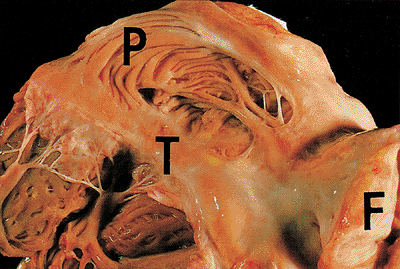 |
Figure 20.11 The right atrium showing the pectinate muscles (P), the crista terminalis (T), and the interatrial septal fat (F). |
Right Atrium
Venous return to the heart occurs via the IVC and SCV into the right atrium. The right atrium forms the right lateral cardiac border and is located anterior to the left atrium and to the right of it. Anteromedially, the right atrial appendage protrudes from the right atrium and overlaps the aortic root (17). The atrium is composed of an auricular appendage, a smooth-walled venous sinus that contains the openings of the cavae and coronary sinus, a septal component, and a portion near the opening of the tricuspid valve (19). Extending between the right sides of the openings of both cavae is a prominent muscular ridge called the terminal crest, which underlies the sulcus terminalis. The wide-based triangular appendage defines the anatomic right atrium. The interior surface of this appendage is trabeculated by muscular bands called pectinate muscles. The portion of the right atrium lateral to the terminal crest is smooth-walled and is derived embryologically from the sinus venosus (Figure 20.11). The right atrial wall measures 2 mm in thickness.
The orifice of the SVC is valveless. The opening of the IVC has an inconstant, rudimentary valve called the eustachean valve. It forms a crescentic fold over the orifice and directed blood toward the foramen ovale and into the left atrium during fetal life. The opening of the coronary sinus has a rudimentary flap of tissue called the thebesian valve; these valves vary greatly in size and may be fenestrated or absent. A Chiari's network, usually appearing as a lacelike veil of tissue, represents the remnant of the right sinus venosus valve and is occasionally found in normal hearts (Figure 20.12). The right atrial chamber is covered by endocardium. In the subendocardium, elastic fibers pass into a typical fenestrated elastic membrane that contains blood vessels, nerves, and branches of the conducting system. In the spaces between the muscle bundles of the atria, the wall is so thin that the connective tissue of the endocardium blends with the epicardium. The epicardial surface of the atrium is rich in nerves and ganglia (Figure 20.13).
Left Atrium
Like the right atrium, this chamber consists of an appendage, a venous component containing the ostia of the four pulmonary veins, a septal component, and a region near the mitral valve orifice. On average, the wall of the left atrium is 3 mm in thickness. The septal surface is smooth. The four pulmonary veins open into the left atrium in the posterior wall. The atrial appendage has a narrow, angulated tubular shape resembling a hockey stick and is lined by pectinate muscles. At its interface with the venous portion of the atrium, it lacks a terminal crest. The endocardial layer is thicker and more opaque than on the right side, due in part to the higher pressures of the pulmonary veins emptying into the atrium. Sometimes a patch of thickened rough
P.533
endocardium is seen on the posterior wall above the mitral valve as a result of mitral regurgitation. This is called MacCallum's patch but is not usually seen in the normal heart. The two left pulmonary veins enter on the posterolateral surface, and the right veins enter on the posteromedial side; there are no true valves at the venous-atrial junction. Small collections of atrial muscle can be found within the walls of the pulmonary veins at this junction and may act as physiologic valves. On occasion, this is the source of isolated or left-sided atrial fibrillation and is treated by ablation techniques.
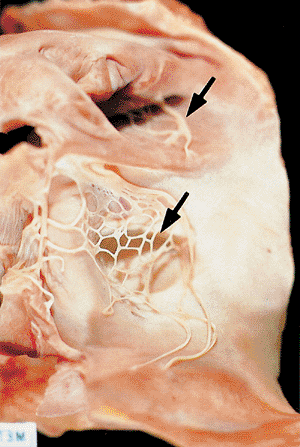 |
Figure 20.12 Picture of the Chiari's network (arrows), or lacelike pattern of the remnants of the thebesian valve, in the inferior vena cava opening into the right atrium. |
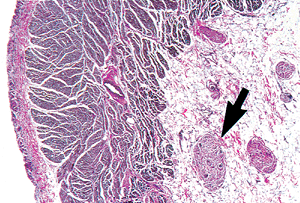 |
Figure 20.13 Section through the right atrium showing the nerves and ganglion cells (arrow) in the overlying subepicardial adipose tissue (elastic van Giesen). |
The atrial septum is smooth on the left side but has a central shallow depression corresponding to the oval fossa. The endocardium of the left atrium is thicker as a result of increased collagen layers, particularly near the openings of the pulmonary veins. Otherwise the histology is similar to the right atrium.
Right Ventricle
The morphologic right ventricle lies anterior to the other heart chambers. It has an inflow portion (sinus), an apical trabecular component, and an outflow portion (infundibulum conus) (20). The outflow tract is separated anatomically from the inflow tract by a muscular arch called the crista superventricularis. The endocardial surface is coarsely trabeculated, particularly in the apica region.
On sectioning, the trabeculations form the inner two-thirds of the ventricular wall. The septal surface is deeply trabeculated with coarse trabeculae carneae and a thick muscular column called the moderator band (Figure 20.14). This moderator band is found in most hearts and connects the distal portion of the septum to the free wall. It terminates in the region of the anterior papillary muscle. Myocyte disarray is a common finding within trabecular muscles and should not be confused with hypertrophic cardiomyopathy. The right bundle branch travels through the muscular ventricular septum and courses down to end in the moderator muscle. The papillary muscles of the right ventricle are relatively constant, with an anterior papillary muscle located on the anterior wall near its junction with the septum and a small posterior papillary muscle arising under the crista superventricularis at the inferior border of the right ventricular outflow tract. In addition, there is an
P.534
inconstant group of posterior papillary muscles that can arise from the diaphragmatic wall of the right ventricle.
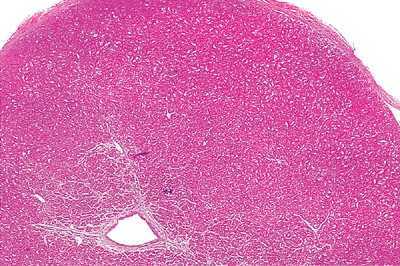 |
Figure 20.14 A section of trabeculae carneae from the normal right ventricle. |
The histology of the right ventricle consists of a thin endocardial layer. The thickness of myocardial wall in normal human adults is about one-third that of the left ventricle and measures up to 5 mm. The endocardium is similar to the other chambers except it has more variability from region to region, with the thickest area found in the septum. The subendocardial space includes a fenestrated elastic membrane and, on occasion, bundles of smooth muscle, particularly in the interventricular septum (Figure 20.7). The interventricular septum also contains blood vessels, nerves, and the left bundle branch of the conduction system. The ventricular free wall has numerous vascular channels consisting of intertrabecular channels that lead into myocardial sinusoids and thebesian veins. Myocardial sinusoids are also found within the trabeculae. Arterioluminal vessels, leading directly from the systemic coronary circulation into the capillary beds, empty into the myocardial sinusoids. The myocardium is richly supplied with small vascular channels that form an intramural circulation (21). There is an extensive web of capillaries that course among the cardiac muscle fibers, are fed by branches of the coronary arteries, and are drained, in part, by the coronary veins. They are also directed to the intramyocardial sinusoids and thence into the lumen of the heart. Deep within the myocardial musculature is found, in addition to the capillary bed, a richly anastomosing network of irregular channels that have been called myocardial sinusoids. These sinusoids receive vessels from the coronary arteries and the capillaries and communicate with coronary veins. The connections between the coronary arteries and the cardiac chambers are called arterioluminal vessels (Figure 20.15). Within the myocardium, variable amounts of adipose tissue can be found, particularly in the free wall portions. When extensive, it is called fatty infiltration of the right ventricle and represents a metaplastic change. It should not be confused with arrhythmogenic right ventricular dysplasia/cardiomyopathy.
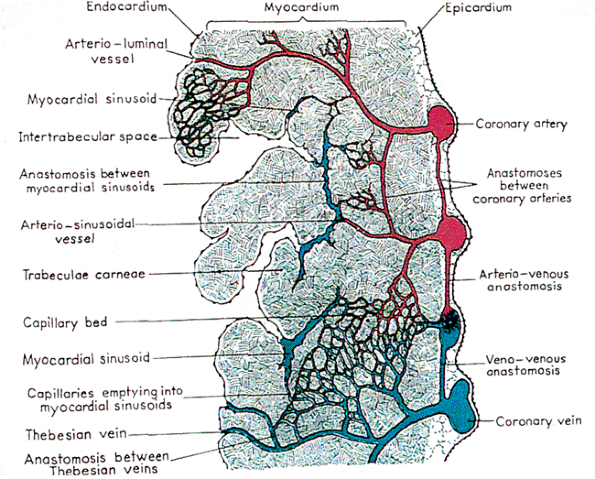 |
Figure 20.15 Diagrammatic representation of the various intramural vascular channels. Reprinted with permission from: Gittenberger-de Groot AC, Bartelings MM, Deruiter MC, Poelmann RE. Basics of cardiac development for the understanding of congenital heart malformations. Pediatr Res 2005;57:169 176. |
Applied Anatomy
The importance of recognizing the presence of myocyte disarray and adipose tissue as common findings within the trabeculated musculature of the right ventricle of the normal adult heart has been discussed. The apical trabecular region is also the site of pacemaker wire placement and endomyocardial biopsy sampling.
Left Ventricle
The left ventricle receives blood from the left atrium during ventricular diastole and ejects blood into the systemic arterial circulation across the aortic valve during ventricular systole. The left ventricle is somewhat bullet-shaped, with the blunt tip directed anteriorly and inferiorly and to the left (22). Like the right ventricle, it is composed of inflow, septal, and outflow components. It lacks the moderator band and septomarginal trabeculations of the right ventricle (19). The left ventricular chamber is surrounded by a thick muscular wall measuring up to 15 mm in thickness (note: the papillary muscles are not included in the measurement). The medial wall of the left ventricle is the interventricular septum, which is shared with the right ventricle. The septum is roughly triangular in shape, with the base of the triangle at the level of the aortic cusps; it is entirely muscular, except for the small membranous septum located just below the right coronary and posterior cusps. The upper third of the septum, or outflow tract, is lined by smooth endocardium. The inferior two-thirds of the septum and the remaining ventricular walls are composed of the trabeculae carneae, which are thinner and less prominent than in the right ventricle. The free wall of the left ventricle is that portion that is exclusive of the septum.
The histology of the left ventricle is similar to that of the right side, although the endocardium is slightly thicker as a result of higher hemodynamic pressures. The small arterioles subadjacent to the endocardium have slightly thicker walls than those of the right ventricle, most likely due to the higher pressure in the left ventricle. The myocardium of the left ventricle is arranged in such a way that it appears to spiral inward from the superficial layers. The superficial layers run at right angles to the layers deeper within the wall. These layers are intimately interdigitated to prevent dissection into lamina structures. The attachment of the muscular layers is from the fibrous skeleton at the base of the heart. The deeper muscle layers of the interventricular septum are composed of the deep bulbospiral muscle in the center of the septum originating from the septal portion
P.535
of the atrioventricular annulus. The fascicles of the deep bulbospiral and sinospiral muscles interdigitate within the muscular interventricular septum. The spiral muscle pulls the base of the ventricle toward the apex during systolic contraction. The blood supply is similar to that of the right ventricle.
Applied Anatomy
Portions of the left ventricular subaortic outflow tract may be submitted as myomectomy specimens in patients with asymmetric hypertrophic cardiomyopathy. Portions of the left ventricular free wall may be removed as aneurysectomy specimens or during VAD placement. The intermediate bulbospiral muscle of the interventricular septum is primarily involved in idiopathic hypertrophic subaortic stenosis (IHSS). Because it lies deep within the septum, the disarray associated with IHSS may not always be seen in the superficial myomectomy specimen.
Cardiac Valves
Semilunar Valves
The semilunar valves consist of the pulmonic and aortic valves. The normal valve circumference for the aortic valve is 5.7 to 7.9 cm for women and 6.0 to 8.5 cm for men (23). The normal pulmonic valve circumference is 5.7 to 7.4 cm for women and 9.2 to 9.9 cm for men (23). Interestingly, when adjusted for body surface area, the valves in women are slightly larger than in men (6). Each semilunar valve consists of three semicircular cusps (24), each of which is attached by its semicircular border, or annulus, to the aortic or pulmonic ring. The three points of lateral attachment of adjacent cusps are the commissures (Figure 20.16). The lines of cusp apposition are not at the free margin but are angulated lines extending from well below the point of attachment in the commissure to just below the midpoint of the free edge. In the aortic valve, these lines (the linea alba) and the central nodules (the noduli arenti) can be seen (24). In the pulmonic valve, these landmarks are less obvious because of the lower right-sided pressures. The lunulae are thin, delicate areas of cusp between the linear alba and the free edge.
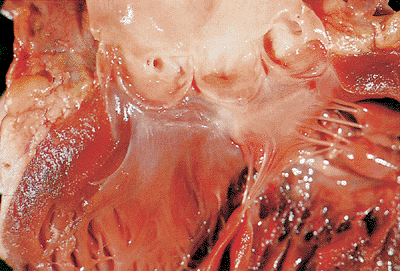 |
Figure 20.16 Aortic valve from normal heart showing the cusps and the coronary ostia. |
The semilunar aortic and pulmonic valves are similar in configuration, except that the aortic cusps are slightly thicker and contain coronary ostia. They are situated at the summit at the outflow tract of their corresponding ventricle, the pulmonic valve being anterior, superior, and slightly to the left of the aortic valve in the normal heart. The cusps, which are often slightly unequal in width, circle the inside of the respective vessel root (e.g., pulmonary artery trunk or aortic root). Behind each cusp, the vessel wall bulges outward, creating a pouchlike dilatation known as the sinus of Valsalva. The portion of the cusp adjacent to the rim is thin and may contain small perforations in the normal situation. The noduli arenti meet in the center and contribute to the support of the leaflets. Because the plane of the aortic valve is oblique with the right posterior side lower than the left anterior side, the origin of the left coronary artery is slightly superior to that of the right coronary artery. The ostia of coronary arteries are located in the upper third of their respective sinuses. The right coronary artery passes anteriorly and to the left. In some hearts, there is a separate ostium in the right coronary sinus for the conus artery, sometimes called the third coronary artery (22).
The histology of the semilunar valves is that of a well-defined multilayered structure. Three distinct layers are recognizable: the fibrosa, the spongiosa, and the ventricularis. The fibrosa is a layer of dense collagen that constitutes the major structural component of the cusp and extends to its free edge (25). The densely packed collagen bundles blend into the collagen of the valvular ring in the region of the commissures. Some fibroblasts are present in this layer, as are some very fine elastic fibers. The spongiosa is subadjacent to the fibrosa and occupies a central position in the thickness of the cusp (Figure 20.17). It is best developed in the basal third of the cusp. It does not extend to the free edge, which is composed only of fibrosa and ventricularis layers. The spongiosa is composed of large amounts of proteoglycans, such as glycoaminoglycans, loosely arranged collagen fibrils, scattered fibroblasts, and mesenchymal cells (25). The ventricularis is subadjacent to the spongiosa and is in direct contact with the endothelial layer of the inflow surface of the cusp (i.e., closest to the ventricular surface). It is distinguished from the fibrosa by its abundance of elastic fibers. This feature is often helpful in orienting and identifying the layers of excised aortic valvular cusps. The surface lining of the cusps consists of a single layer of endothelial cells. The surface topography of the aortic cusp varies according to its state of stress. The bundles are wavy and the inflow surface is smoother in the stressed state and rougher in the relaxed state (25).
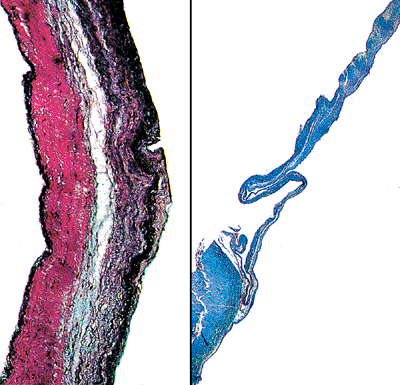 |
Figure 20.17 High- (A) and low-power (B) views of a section of the aortic cusp showing the three distinct layers as described in the text. |
P.536
Atrioventricular Valves
The atrioventricular valves consist of the mitral valve and the tricuspid valve. The normal circumference of the tricuspid valve is 10 to 11.1 cm in women and 11.2 to 11.8 cm in men (23). The normal circumference of the mitral valve is 8.2 to 9.1 cm in women and 9.2 to 9.9 cm in men (23). The valvular apparatus is made up of the annulus, commissures, leaflets, chordae tendineae, and papillary muscles. The annulus is composed of a ring of circumferentially oriented collagen and elastic fibers with extensions into the ventricle and atrium.
The atrioventricular valves have four histologic layers (Figure 20.18). The collagen bundles of the annulus spread down into the cusp of the mitral valve and are known as the fibrosa. They continue into the chordae tendineae and finally spread out into a network that covers the tip of the papillary muscles (Figure 20.19). Adjacent to the fibrosa layer on the ventricular side of the valve is the ventricularis. The ventricularis contains many elastic fibers and is covered by endothelium. Some of the elastic fibers extend into the chordae tendineae; but, in general, this layer is incomplete, as it does not extend to the free edge of the leaflet. The spongiosa is situated on the atrial side of the fibrosa layer. Like the semilunar valves, it has a rich abundance of glycosaminoglycans and a few elastic fibers, collagen fibrils, and connective tissue cells (25). This layer, along with the fibrosa layer, extends through the entire length of the leaflet. The spongiosa in the anterior and posterior mitral leaflets may contain cardiac myocytes that are a direct extension of the left atrial myocardium (Figure 20.18). In the anterior mitral leaflet, this layer extends into the middle third, whereas in the posterior leaflet it only extends into its proximal third. Neural elements and lymphatics can be found in the leaflets. The atrial aspect of the spongiosa (i.e., layer closest to the atrium) is covered by the auricularis,
P.537
which in turn has an endothelial lining. The auricularis contains collagen and elastic fibers and smooth muscle cells. It is prominent near the annulus but thins out in the distal third of the leaflet such that the most distal aspect of the valves is composed only of spongiosa and fibrosa. The endothelial cells on the atrial aspect of the atrioventricular valves are plump and have irregular nuclei compared to the flatter endothelial cells on the ventricular aspect.
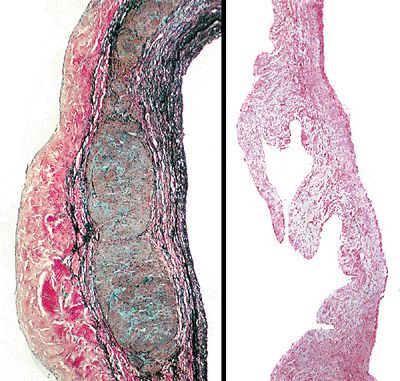 |
Figure 20.18 High- (A) and low-power (B) views of a section of the mitral leaflet showing the three distinct layers with muscle in the central portion near the base of the valve. |
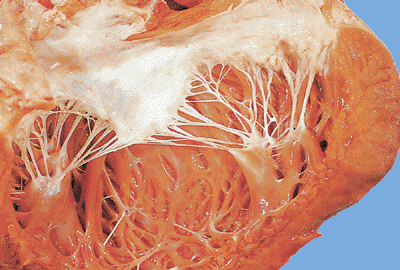 |
Figure 20.19 A normal mitral valve with chordae tendineae inserting into the papillary muscles. |
The architecture of the tricuspid valve apparatus and the layered arrangements of its leaflets are similar to those in the mitral valve; however, the individual layers are thinner in the tricuspid valve. Cardiac muscle bundles insert fairly low into the base of the tricuspid leaflets but do not extend into the leaflet substance. In the posterior and septal leaflets, the auricularis is thicker and contains more abundant smooth muscle cells.
Chordae Tendineae
The chordae tendineae in the normal state are thin fibrous cords that emanate in a fanlike manner from the broad leaflets of the atrioventricular valves and insert into the papillary muscles (Figure 20.19). The central cores of the chordae are composed of longitudinally oriented collagen fibrils. The core is surrounded more peripherally by loosely arranged collagen fibrils and elastic fibers and is embedded in proteoglycan-rich matrix (25) (Figure 20.20). Some chordae contain a small central core of muscle known as chordae muscularis (Figure 20.21), although others contain blood vessels and collagen in variable amounts and appear fleshier in color (Figure 20.22). The endothelial cells on the chordae resemble the flattened ovoid nuclei of the ventricular surface of the leaflets.
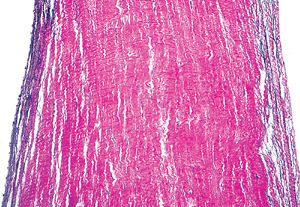 |
Figure 20.20 Longitudinal section of chordae tendineae showing the relationships of elastic fibers on the surface and collagen in the center (elastic van Giesen). |
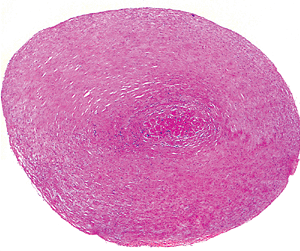 |
Figure 20.21 Transverse section of chordae tendineae showing central core of muscle (H&E). |
Applied Anatomy of Intracardiac Valves
Semilunar and atrioventricular valves are frequently encountered in the surgical pathology laboratory. Indications for valvular replacement or repair include a variety of congenital, infectious, inflammatory, degenerative, and paraneoplastic causes. In many cases, the chordae and portions of papillary muscle may be attached. In the
P.538
setting of myocardial infarction, infection, or valvular prolapse, ruptured papillary muscles may become surgical specimens. Lambl's excrescences and fibrous nodules are papillary projections along the lines of closure or free edge of the valve, respectively.
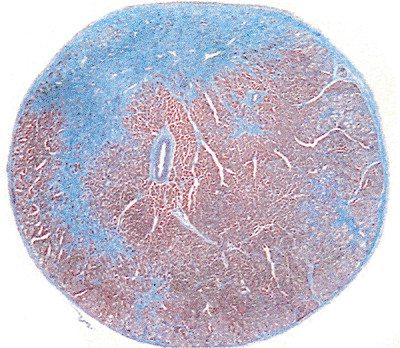 |
Figure 20.22 Transverse section of a muscular chorda showing fibrous tissue, muscle, and small vessel within the chorda (Masson's trichrome). |
Aging Changes of Intracardiac Valves
With age, all the cardiac valves become thicker, more opaque, and less pliable. The increase in collagen content may account for the loss of plasticity, and calcifications may occur. The posterior leaflet of the mitral valve often shows yellow atheromatous alteration. The mitral valve annulus may become calcified with age. The valvular circumferences increase with age (6).
Papillary Muscles
The two papillary muscles of the right ventricle (anterior and conal) are relatively constant. There is also a group of inconstant posterior papillary muscles on the inferior wall. In the left ventricle there are two constant papillary muscles: the anterior and posterior. The papillary muscles receive the chordae. They are variable in shape and width and on occasion may have multiple heads (Figure 20.19).
The histology of the papillary muscles includes the fibrous cap, into which the chordae insert (Figure 20.23). The small arteries and arterioles in the papillary muscle are notable for their wall thickness and irregularity in comparison to other intracardiac small vessels (Figure 20.24). The myocardium and the endocardial covering are similar to their counterparts described elsewhere. In marked ventricular dilatation, the papillary muscles may become thinned and flattened.
Conduction System
Myocardial fibers are delineated along two functional pathways in humans: (a) contractile fibers and (b) myocardial fibers specialized for the initiation and propagation of an impulse for contraction. The conduction system is recognized to be myogenic in origin, with nerves playing only a subsidiary controlling function.
For most pathologists, examining the conduction system is often regarded as a daunting exercise. This is due in large part to the fact that the number of cases requiring detailed morphologic analysis is infrequent and is often limited to specific requests from clinicians. In addition to detailing the microscopic features of the sinoatrial and atrioventricular nodes, we will present our practical approach to the dissection of these structures. We recommend careful attention to key anatomic landmarks to ensure successful retrieval of these structures.
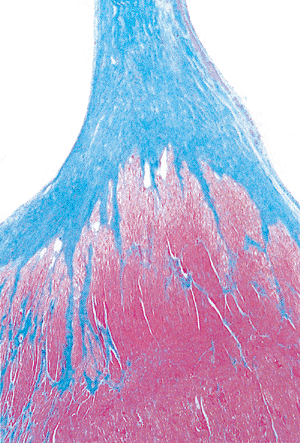 |
Figure 20.23 Longitudinal section of papillary muscle showing the fibrous cap of the insertion of the chordae tendineae (Masson's trichrome). |
Sinoatrial Node
The sinoatrial (SA) node has the highest intrinsic rate and is recognized as the primary pacemaker of the heart. This node
P.539
is situated within the terminal groove at the junction of the SVC and the lateral border of the right atrium (Figure 20.25). Its position is constant and is marked by the apex of the crest of the atrial appendage. It is ovoid or cigar-shaped in most hearts, but rare cases of horseshoe-shaped SA nodes are reported. Removal of a rectangular block of tissue that includes the distal SVC and atrium on either side of the terminal groove is recommended. Serial sectioning in a longitudinal plane parallel to the terminal groove at 2 mm intervals is recommended, and all the tissue slices can be accommodated in 2 tissue cassettes. Grossly identifying the SA nodal artery is also helpful in procuring the node (Figure 20.25). This is a branch of the right coronary artery that is found in slightly more than half of the general population. Microscopically, the node is arranged around a central artery adjacent to epicardial adipose tissue. The node is composed of dense connective tissue within which the small muscle fibers are embedded. The muscle fibers contain sparse myofibrils, the striations are not prominent, and the whole mass has a pseudosyncytial appearance. Connective tissue stains such as Masson's trichrome and elastic van Gieson stains highlight the abundant collagen and elastic fibers, respectively (Figure 20.25). Abundant nerve fibers run into the node. The exact pathway(s) carrying the electrical impulse from the SA node to the atrioventricular node remains controversial. Some investigators think that several specialized bundles of conducting system cells (e.g., anterior, middle and posterior internodal tracts) conduct the impulse around the atrium. Others argue that the arrangement of myocardial fibers within the atrium and interatrial septum serves to propagate the impulse (16).
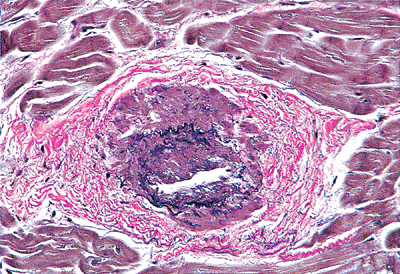 |
Figure 20.24 Section of abnormally thickened arteriole within a papillary muscle (elastic van Giesen). |
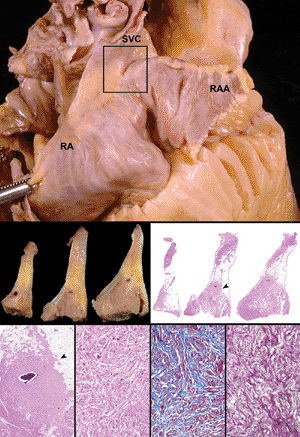 |
Figure 20.25 The sinoatrial node. A. The location of the sinoatrial node within the terminal groove at the junction of the superior vena cava (SVC) and crest of the atrial appendage (box). B. Macroscopic and low-power magnification of serial sections of nodal tissue. Note the nodal artery that is adjacent to the sinoctrial node. C. High-power magnification of the sinoatrial node showing the specialized fibers embedded within collagen and elastic tissue (H&E, trichrome, and elastic van Giesen stains). |
Atrioventricular Node
In our experience a heart opened along the lines of flow provides the optimum exposure for dissecting the atrioventricular (AV) node. From a right-sided approach the interventricular septum is oriented with the tip of the apex pointing downward. The important landmarks include the oval fossa, ostium of the coronary sinus, and tricuspid valve annulus and leaflets. The AV node lies within the subendocardial tissues on the right side of the interatrial septum just anterior to the opening of the coronary sinus, posterior to the membranous interventricular septum, and above the tricuspid valve annulus within the triangle of Koch (26) (Figure 20.26). A rectangular block of tissue, beginning with a vertical incision adjacent to the ostium of the coronary sinus and extending 1 to 2 cm below the annulus, is removed. After careful trimming of valvular structures, the block will contain components of the tricuspid valve (septal leaflet) and mitral and aortic valves (Figure 20.26). Serial sectioning at 2 to 3 mm intervals and sequential placement in tissue cassettes yields a total of 8 to 10 cassettes.
In histologic sections, the AV node is flattened against the central fibrous body and is composed of a network of muscle fibers, with the superficial zone having fibers arranged in a parallel manner. These specialized fibers retain their intercalated disks and striations but are characterized by their pale eosinophilic appearance (Figure 20.26). A small AV nodal artery is often identified adjacent to the AV node. At the anterior end of the AV node, the muscle fibers become arranged in parallel lines to form the main bundle of His or penetrating AV bundle. To reach the ventricle, the AV bundle pierces the central fibrous body and runs forward on the upper margin of the muscular ventricular septum. This penetrating portion of the main bundle is surrounded by dense connective tissue and anatomically is closely related to the aortic and mitral valve rings (Figure 20.26). Connective tissue stains can aid in the localization of the nodal tissue. The
P.540
fibers of the main bundle are arranged in parallel. The penetrating AV bundle terminates as the left and right bundle branches. The left fascicle runs downward over the endocardial surface of the interventricular septum to the base of the anterior papillary muscle, and the right fascicle ends in the moderator band of the right ventricle. Direct connection of both bundle branches to a complex ramifying system of subendocardial conduction fibers can be demonstrated in mammalian hearts. Light microscopy shows the fibers in the bundle of His and the conduction bundles to be small and contain few myofibrils (Figure 20.27).
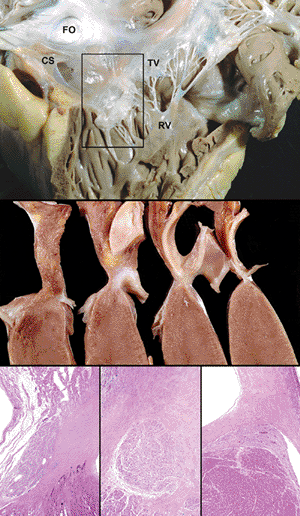 |
Figure 20.26 The atrioventricular nodal apparatus. A. The location of the AV node (box) viewed from the right ventricle. Important landmarks include the coronary sinus (CS), tricuspid valve annulus and leaflet (TV), and fossa ovale (FO). B. Serial sections of the rectangular block of tissue show the relationship of the nodal tissue to the tricuspid valve, mitral valve, atrioventricular valve, and fibrous skeleton of the heart. C. The atrioventricular (AV) node (left), AV penetrating bundle (middle) and AV bundle with fascicle (right) are shown. |
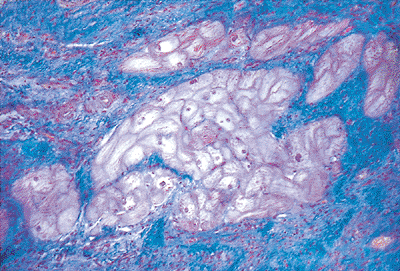 |
Figure 20.27 Section showing the pale cells of the mammalian conducting system. These cells contain glycogen and only sparse myofibrils (Masson's trichrome). |
Aging Changes in the Human Conduction System
With advancing age, the SA node displays progressive increase in fibrous tissue while the AV node remains relatively unchanged. Similarly, fibrous tissue increases in the upper portion of the interventricular septum. These changes are associated with a loss of conduction fibers in the region of the left bundle branch. Up to 50% of the left bundle origin may be lost in people over 60 years of age (27).
Cardiac Innervation
The nerve supply of the heart is autonomic, including both the sympathetic and parasympathetic supply via both the efferent and afferent fibers. Histologically, large nerves can be seen in the epicardium and adjacent to the coronary blood vessels. Small nerves within the myocardium are hard to identify unless special stains are used. Myocardial nerves are best viewed using electron microscopic examination, by which the autonomic nerves can be distinguished. Cardiac ganglia (parasympathetic) can be found over the surface of the atria and in the AV groove (Figure 20.13).
Autonomic Nerves
Axonal varicosities occur at irregular intervals along autonomic fibers, and their morphology is considered useful in determining whether the nerve is adrenergic or cholinergic (25). In cholinergic nerves, the varicosities contain accumulations of agranular vesicles and a few mitochondria. In adrenergic nerves, the varicosities contain vesicles rich in electron-dense cores. Each of these cores is separated from the limiting membrane of the vesicle by an electron-lucent zone (Figure 20.28). Presumptive sensory nerve terminals
P.541
have large diameters and contain numerous mitochondria. They are located in perivascular regions and are surrounded by Schwann cells. A given Schwann cell may enclose adrenergic and cholinergic axons together with sensory axons (25). Autonomic ganglia are found in the subepicardial tissue of the atria and atria appendages and at the root of the great vessels, along the interatrial and AV grooves in the atrial septum and in the vicinity of the SA and AV nodes. Large nerves can be seen in the subepicardial layer adjacent to the epicardial coronary arteries.
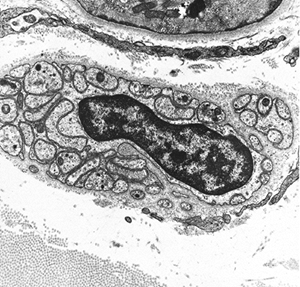 |
Figure 20.28 Electron micrograph of sympathetic nerve showing dense-core granules in the myocardium (original magnification 22,500). |
 |
Figure 20.29 Electron micrograph showing an intramyocardial lymphatic with thin walls and no basement membrane (original magnification 20,000). |
Lymphatics
There are two networks of lymphatics in the heart: (a) in the endocardium and (b) in the epicardium. The route of drainage of the endocardial network is through channels in the myocardium into the epicardial lymphatics. The epicardial meshwork of channels, containing many valves, drains toward the AV sulcus by means of several longitudinal channels that run for the most part parallel to the coronary veins in the anterior and posterior longitudinal sulci of the ventricles (25). Lymphatics leave the pericardial cavity to empty into one of the pulmonary hilar lymph nodes and join the lymphatic drainage system of the mediastinum or into the thoracic duct. Lymphatics are also found in the myocardial valves and lie within the grooves of the coronary blood vessels.
The lymph capillaries and larger lymphatic vessels accompany blood vessels in the myocardial interstitium. The walls of the myocardial lymphatics consist of extremely thin endothelial cells, the nuclei of which bulge into the lumen (Figure 20.29). In contrast to endothelial cells of blood capillaries, those of the lymphatic capillaries do not have a well-defined external basal lamina. The endothelial cells of lymphatic capillaries may have Weibel-Palade bodies and transplant vesicles (25). The larger lymphatics are confined to the outer third of the myocardial wall and occasionally contain valves. These flaplike structures contain a core of collagen embedded in microfibrils and are covered by endothelium.
Small Intramural Coronary Arteries
The structure of the intramural coronary arteries and the larger coronary arteries is similar and consists of the endothelium, smooth muscle, and adventitia (Figure 20.30).
P.542
These smallest muscular arteries contain three or four layers of smooth muscle. Arterioles have flat, elongated endothelial cells that do not protrude into the lumen. Their internal elastic lamina is discontinuous. Metarterioles are also known as precapillary sphincters. The endothelial cells in metarterioles have numerous surface projections that bulge into the lumen (25). Although the medial smooth muscles form a single discontinuous layer, it gradually disappears as capillaries begin. Capillaries are distinguished by the fact that their walls are composed of only a single layer of endothelial cells. They do not have smooth muscle cells but may have closely associated pericytes (25). Capillary endothelial cells may have microvilli and cytoplasmic processes (filopodia). The myocardium has a rich network of capillaries. These branches undergo anastomosis and eventually become thin-walled venules measuring up to 100 m in diameter.
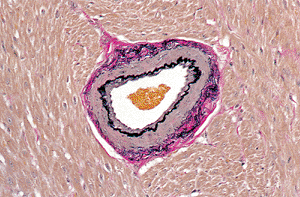 |
Figure 20.30 Transverse section of intramyocardial arteriole (elastic van Giesen). |
Veins and Venules
Venules have thin, flat endothelial cells and characteristically contain a large amount of connective tissue in the vicinity of their external surface; they contain collagen fibrils that approach the endothelial layer and are anchored on its outer surface (25). Venules gradually increase in size to become veins. Veins have larger lumens but thinner walls than their arterial counterparts.
Veins have three layers: the intima, media, and adventitia. The intima is thin, lacks smooth muscle cells and has a poorly defined internal elastic lamina. The media is also thin and contains few smooth muscle cells and elastic fibers. The adventitia is thick with abundant collagen and elastic fibers. Cardiac veins drain blood into either the coronary sinus or directly into the chambers (thebesian veins).
The Endomyocardial Biopsy
The transvenous endomyocardial biopsy is currently utilized for the diagnosis of allograft rejection and a variety of inflammatory, metabolic, and neoplastic conditions that affect the heart. Originally introduced in the early 1960s, the bioptome and the technique have undergone modifications that now permit clinicians the opportunity to obtain cardiac tissue in a safe outpatient setting (28). The right internal jugular vein or femoral vein approaches are commonly used. Complications are uncommon and include local problems (such as hematomas and nerve injury) and cardiac problems (such as arrhythmias, tricuspid valve apparatus damage, and ventricular perforation).
Tissue Handling and Processing
Proper tissue procurement and handling are essential for optimal diagnostic evaluation (29). Biopsy specimens should be gently extracted from the bioptome with a needle tip to limit crush artifactual distortion. The clinical indications for the biopsy determine, in large part, the method of tissue handling. For example, for standard light microscopy, the tissue should immediately be placed in a standard fixative such as 10% neutral buffered formalin. To demonstrate the type of amyloid fibril in cardiac amyloidosis by immunofluorescence (e.g., AL, AA, or transthyretin), one or two pieces should be received in saline or Zeus medium and then snap frozen in a plastic Beem capsule containing an embedding medium. The diagnosis of chronic anthracycline cardiotoxicity requires that all the biopsy pieces (minimum of 3 5 pieces) be submitted in fixative for transmission electron microscopy (e.g., 2.5% glutaraldehyde with 2% paraformaldehyde in 0.1M sodium cacodylate buffer, pH 7.2).
For routine diagnostic evaluation, overnight processing and paraffin embedding are sufficient. For emergent cases, a 90-minute rapid ( ultra ) processing cycle is available, and microscopic slides can be prepared within three to four hours. All the biopsy pieces should be embedded in the same block. We recommend that a minimum of three slides are prepared, with each sectioned at 4 to 5 m thickness from various depths within the paraffin block. Multiple fragments, or ribbons, are placed on each slide.
We routinely stain with hematoxylin and eosin (H&E) and use stains such as Masson's trichrome to confirm the presence of myocyte damage or fibrosis, Congo red stain for amyloid fibrils, and the Prussian blue stain for iron deposition. Immunohistochemical, immunofluorescent, and molecular studies are utilized for specific indications. Paraffin section immunohistochemistry is used to evaluate for the presence of infectious myocarditis (e.g., cytomegalovirus (CMV) or toxoplasmic myocarditis), posttransplant lymphoproliferative disorders (PTLD) [B-cell clonality, Epstein-Barr virus (EBV) latent membrane proteins, anomalous coexpression of B-cell and T-cell antigens] or acute antibody mediated rejection (intravascular collections of CD68+ histiocytes and deposition of C4d on the microvasculature). In situ hybridization is helpful to demonstrate the presence of EBV or other viral genome or light chain restriction in PTLD.
Biopsy Limitations and Tissue Artifacts
Sampling error in the diagnosis of rejection, myocarditis, and infection remains a major consideration in the clinical management of patients and the evaluation of new noninvasive diagnostic modalities. In general, the false-negative rate is low, particularly when four or more pieces of tissue are submitted. The issue of how many lymphocytes are normally found in the myocardium has been addressed. In an endomyocardial biopsy study, the mean number of lymphocytes is fewer than 5.0 per high-power field (30).
A variety of artifacts occur in endomyocardial biopsy specimens that may mimic pathologic lesions. The surgical pathologist must be aware of these patterns to avoid a
P.543
misdiagnosis that could lead to unnecessary therapeutic interventions. These have been reviewed in detail in a recent publication and only selected topics will be briefly reviewed (31). The most common biopsy artifact is the presence of contraction bands in myocytes (Figure 20.31A). They are identical to the linear bands observed in acute ischemic necrosis and catecholamine ( pressor ) effect. These changes are induced by the biopsy procedure itself and can be diminished by using fixatives stored at room temperature. In ischemic injury, the nuclei of surrounding myocytes are usually pyknotic (Figure 20.31B), while in artifactually induced contraction bands, the nuclei remain normal in appearance.
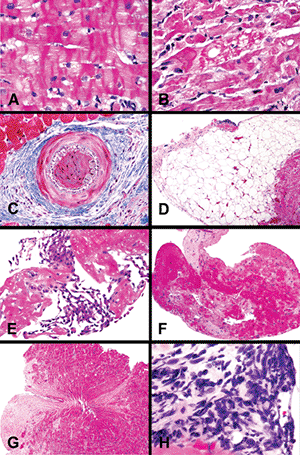 |
Figure 20.31 Artifacts of endomyocardial biopsy specimens: A. Contraction band artifact (H&E); note the normal appearance of myocyte nuclei. B. Contraction band necrosis with hyperchromatic pyknotic nuclei and eosinophilic cytoplasm. The changes are contrasted with the common contraction band artifact in A. C. Telescoping of intramyocardial is highlighted by a trichrome stain. D. Intramyocardial adipose tissue. The presence of fat does not imply epicardial localization or perforation. E. Mesothelial cells admixed with fibrin indicative of ventricular perforation. F. Thrombus without attached myocardial tissue. G. Bioptome-induced Victorian waistband artifact. H. Crush artifactual distortion of cells. |
Another frequent artifact is intussusception, or telescoping, of small arteries that has been confused with luminal occlusion by thrombus and transplant-related arteriosclerosis. Connective tissue stains such as Masson's trichrome or elastic van Gieson highlight the internal elastic membranes of both vessel segments (Figure 20.31C). Intramyocardial accumulations of mature adipose tissue can simulate epicardial tissue, especially if associated with vessels of relatively large caliber (Figure 20.31D). Both can be found in the right ventricular apical region, and adipose tissue is found not uncommonly in woman and elderly patients. This should not be confused with arrhythmogenic right ventricular cardiomyopathy/dysplasia; clinical-pathologic correlation is essential for this purpose.
Ventricular perforation is identified by the presence of mesothelial cells (Figure 20.31E). Accumulations of fresh platelet- and fibrin-rich thrombus may be identified along the endocardial surface of biopsy fragments (Figure 20.31F). These form as a result of the repetitive placement of the bioptome along the endocardial surface and do not indicate chronic mural thrombi. A number of patterns of bioptome-induced tissue distortion or crush artifact can be observed in biopsy samples. The hour-glass or Victorian waistband effect is caused by central constriction of the tissue by the bioptome mechanism (Figure 20.31G). A more problematic artifact is the smearing of cytoplasmic and nuclear components of cells that yields strands of basophilic material (Figure 20.31H). In this setting, it may not be possible to distinguish the cell types (lymphocytes, endothelial cells, histiocytes, myocytes), and we do not attempt to evaluate these foci for allograft rejection or myocarditis. In some cases, procurement of additional leveled H&E-stained sections can provide less distorted foci in the deeper aspects of the biopsy sample. In our experience, immunohistochemical stains have not been consistent or helpful.
Summary
Because of the structural-functional nature of cardiac disease, the surgical pathologist should have a working knowledge of both anatomy and histology. Moreover, the alterations produced by the endomyocardial biopsy and the bioptome require familiarity with the myriad of tissue artifacts. With a practical understanding of these points, the evaluation of specimens ranging from endomyocardial samples to explanted hearts will be enhanced and the diagnostic information to clinicians more precise.
References
1. Hertz MI, Boucek, MM, Deng MC, et al. Scientific Registry of the International Society for Heart and Lung Transplantation: introduction to the 2005 annual reports. J Heart Lung Transplant 2005;24:939 944.
2. Boucek MM, Edwards LB, Keck BM, Trulock EP, Taylor DO, Hertz MI. Registry of the International Society for Heart and Lung Transplantation: eighth official pediatric report 2005. J Heart Lung Transplant 2005;24:968 982.
3. Scholz DG, Kitzman DW, Hagen PT, Ilstrup DM, Edwards WD. Age-related changes in normal human hearts during the first 10 decades of life. Part I. (Growth): a quantitative anatomic study of 200 specimens from subjects from birth to 19 years old. Mayo Clin Proc 1988;63:126 136.
4. Ludwig J, ed. Handbook of Autopsy Practice. 3rd ed. Totowa, NJ: Humana Press; 2002.
5. Hudson R. Structure and function of the heart. In: Hudson R, ed. Cardiovascular Pathology. Vol 1. London: Edward Arnold; 1965:12 23.
6. Kitzman DW, Scholz DG, Hagen PT, Ilstrup DM, Edwards WD. Age-related changes in normal human hearts during the first 10 decades of life. Part II. (Maturity): a quantitative anatomic study of 765 specimens from subjects 20 to 99 years old. Mayo Clin Proc 1988;63:137 146.
7. Hutchins GM, Anaya OA. Measurements of cardiac size, chamber volumes and valve orifices at autopsy. Johns Hopkins Med J 1973;133:96 106.
8. Sheaff MT, Hopster DJ. Organ dissection cardiovascular system. In: Sheaff MT, Hopster DJ. Post Mortem Technique Handbook. London: Springer; 2001.
9. Silver MM, Silver MD. Examination of the heart and of cardiovascular specimens in surgical pathology. In: Silver MD, Gotlieb AI, Schoen FJ, eds. Cardiovascular Pathology. New York: Churchill Livingstone; 2001:1 29.
10. Edwards WD. Applied anatomy of the heart. In: Brandenberg RO, Fuster V, Guiliani ER, McGoon ER eds. Cardiology: Fundamentals and Practice. Chicago: Year Book Medical; 1987:47 112.
11. Cook AC, Yates RW, Anderson RH. Normal and abnormal fetal cardiac anatomy. Prenat Diagn 2004;24:1032 1048.
12. Foley A, Mercola M. Heart induction: embryology to cardiomyocyte regeneration. Trends Cardiovasc Med 2004;14:121 125.
13. Gittenberger-de Groot AC, Bartelings MM, Deruiter MC, Poelmann RE. Basics of cardiac development for the understanding of congenital heart malformations. Pediatr Res 2005;57:169 176.
14. Pandur P. What does it take to make a heart? Biol Cell 2005;97:197 210.
15. Burke A, Virmani R. Tumors of the heart and great vessels. In: Atlas of Tumor Pathology. 3rd series, fascicle 16. Washington, DC: Armed Forces Institute of Pathology; 1995:127 170.
16. Anderson RH, Becker AE. The Heart: Structure in Health and Disease. London: Gower Medical Publishing; 1992.
17. Malouf JF, Edwards WD, Tajik AJ, Seward JB. Functional anatomy of the heart. In: Fuster V, Alexander RW, O'Rouke RA, eds. Hurst's the Heart. 11th ed. New York: McGraw-Hill; 2004:45 86.
18. Sweeney LJ, Rosenquist GC. The normal anatomy of the atrial septum in the human heart. Am Heart J 1979;98:194 199.
19. Sheppard M, Davies MJ. Practical Cardiovascular Pathology. London: Arnold Publishers; 1998.
20. Davies MJ. Introduction to normal cardiac anatomy. In: Davies MJ, Mann JM. The Cardiovascular System. Part B: Acquired Diseases of the Heart. New York: Churchill Livingstone; 1995:1-6.
P.544
21. Barry A, Patten B. The structure of the adult heart. In: Gould SE, ed. Pathology of the Heart and Blood Vessels. Springfield, IL: Charles C Thomas; 1968:104-105.
22. James TN, Sherf L, Schlant RC, Silverman ME. Anatomy of the heart. In: Hurst JW, Logue RB, Rackley CE, et al., eds. The Heart. 5th ed. New York: McGraw-Hill; 1982:22 74.
23. Silver MM, Freedom RM. Gross examination and structure of the heart. In: Silver MD, ed. Cardiovascular Pathology. 2nd ed. Vol 1. New York: Churchill Livingstone; 1991:1 42.
24. Davies MJ, Pomerance A, Lamb D. Techniques in examination and anatomy of the heart. In: Pomerance A, Davies MJ, eds. Pathology of the Heart. Oxford: Blackwell Scientific; 1975:1 48.
25. Ferrans VJ, Rodr guez ER. Ultrastructure of the normal heart. In: Silver MD, ed. Cardiovascular Pathology. 2nd ed. New York: Churchill Livingstone; 1991:43-101.
26. Edwards WD. Cardiovascular system. In: Ludwig J, ed. Handbook of Autopsy Practice. 3rd ed. Totowa, NJ: Humana Press; 2002:21-44.
27. Davies MJ, Anderson RH. The pathology of the conduction system. In: Pomerance A, Davies MJ, eds. The Pathology of the Heart. Oxford: Blackwell Scientific; 1975:367 412.
28. Baughman KL. History and current techniques of endomyocardial biopsy. In: Baumgartner WA, Reitz B, Kasper E, Theodore J, eds. Heart and Lung Transplantation. 2nd ed. Philadelphia: WB Saunders; 2002:267-281.
29. Berry GJ, Billingham ME. The pathology of human cardiac transplantation. In: Baumgartner WA, Reitz B, Kasper E, Theodore J, eds. Heart and Lung Transplantation. Philadelphia: WB Saunders; 2002:286-306.
30. Edwards WD, Holmes DR Jr, Reeder GS. Diagnosis of active lymphocytic myocarditis by endomyocardial biopsy: quantitative criteria for light microscopy. Mayo Clin Proc 1982;57:419 425.
31. Hauck AJ, Edwards WD. Histopathologic examination of tissues obtained by endomyocardial biopsy. In: Fowles RE, ed. Cardiac Biopsy. Mount Kisco, NY: Futura Publishing; 1992:95 153.