32 - Spleen
Editors: Mills, Stacey E.
Title: Histology for Pathologists, 3rd Edition
Copyright 2007 Lippincott Williams & Wilkins
> Table of Contents > X - Female Genital System > 40 - Vagina
40
Vagina
Stanley J. Robboy
Rex C. Bentley
Introduction
Tissue from the vagina is infrequently examined via biopsy. Excluding the vaginal cuff removed for cervical disease, most biopsies and surgical operations are for infection, small intramural growths, intrauterine exposure to diethylstilbestrol (DES), or, in older women, squamous cell cancer and its precursors. This chapter addresses the gross, microscopic, and ultrastructural anatomy of the normal vagina. The embryologic discussion focuses on developmental perturbations, which provide insights into normal and microscopic anatomy.
Embryologic Changes
The paired m llerian (paramesonephric) ducts appear about the thirty-seventh day postconception as funnel-shaped openings of celomic epithelium (1,2). These develop into paired, undifferentiated tubes that later grow caudally, using the already formed wolffian (mesonephric) ducts as a guidewire to reach the level of the future hymen (Figure 40.1). The absence of this occurrence, the incidence of which is about 1 in 5,000 newborn girls, results in the Mayer-Rokitansky-Kuster-Hauser syndrome (absence of the vagina and cervix with usually only the remnants of the uterine horns) or complete absence of the m llerian derivatives [vagina, uterus, and fallopian tubes] (3).
At about day 54, the paired m llerian ducts fuse, becoming a straight uterovaginal canal (primordia of uterine corpus, cervix, and vagina), the lining of which is an immature columnar (M llerian) epithelium (Figure 40.2) (4). The above changes occur in both female and male embryos. If the fetus is a male, the indifferent gonads become anatomically distinct testes at around day 44. The testis is important for two products it makes. One, m llerian-inhibiting substance (MIS), affects the future of the m llerian ducts. The other, testosterone, affects the future of the wolffian ducts. Shortly after the testes become distinct, Sertoli cells initiate MIS production, a protein in the large transforming growth factor- family, in amounts effective to cause the m llerian ducts to regress through a process of programmed cell death (5,6). If the embryo is female, testes do not develop. Because there is then no MIS, the m llerian ducts are not inhibited from developing and thus grow without impedance to become the uterine tubes, uterus, and vagina.
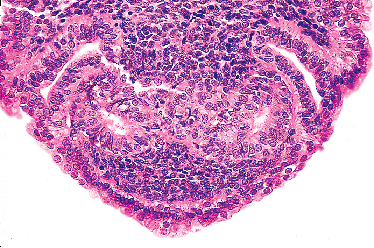 |
Figure 40.1 Region of urogenital sinus disclosing the tips of two central m llerian ducts that have grown down the (outer) paired wolffian ducts (circa day 54). The cytologic features of the cells comprising both types of duct are indistinguishable on light microscopy at early stages of development. Reprinted with permission from: Robboy SJ, Ellington KS. Pathology of the Female Genital Tract in Kodachrome Slides. 2nd ed. Durham, NC: Gyn-Path Associates; 1996. |
P.1000
In contradistinction to MIS, which acts as an inhibitor, testosterone stimulates and is required to promote wolffian duct growth. In the male, the critical period for testosterone stimulation begins early in the tenth week (circa day 71) and causes the embryonic wolffian ducts to differentiate into epididymis, seminal vesicle, and ductus (vas) deferens. If there are no testes (as in the female) and testosterone stimulation has not occurred by the close of the critical window (circa day 84), the wolffian ducts wither and become vestigial remnants, which in the adult are found deep in the vaginal wall.
Until near the end of week 10 (to about day 66) the uterovaginal canal, with its solid tip already in contact with the urogenital sinus, continues to elongate caudally, still remaining as a simple tube lined by columnar epithelium (Figure 40.2). Beginning in the eleventh week, the epithelium stratifies, becoming several layers thick. Under one theory, the squamous cells are believed to derive from the urogenital sinus, invading the common tube from below and growing up the muscular scaffold to completely replace the m llerian epithelium to the level of the external cervical os (7). The transition from m llerian to squamous epithelium occurs at about the time when nuclear estrogen receptors appear in the vaginal stroma (8,9).
Stratification of the squamous epithelium lining the uterovaginal canal heralds formation of the so-called vaginal plate (circa the eleventh week). The proliferation progressively occludes the canal beginning caudally and extending cranially. Studies with latex-injected specimens indicate that even the lateral wings of the vaginal plate may become occluded, but a central lumen persists, especially in the upper vagina.
During the thirteenth week (91 days), cervical glands develop; they exhibit a wavy architectural appearance but cytologically are minimally differentiated. By the fourteenth week, the caudal vagina increases markedly in size (Figure 40.3). During the fifteenth week, the solid epithelial anlage of the anterior and posterior vaginal fornices appear. Starting with the sixteenth week, the squamous epithelium lining the vagina and the exocervix begins to mature, thus resembling the lining of the adult vagina. The epithelium thickens and glycogenates, features most likely related to increased maternal and hence fetal estrogen levels. As the cells mature, they lose cellular adhesiveness and desquamate, heralding the canalization of the vaginal plate and thus the onset of the final gross adult structure of the vagina. By the eighteenth to twentieth week, the development of the vagina is complete.
Why columnar epithelium with an embryonic appearance should initially line the m llerian system and later be replaced by squamous epithelium remains of teleologic interest. The answer to the mechanism may lie in the vaginal wall stroma. Prior work in the mouse has shown that differentiation of the lower genital tract epithelium is dependent on the stroma on which it grows (10). For example, uterine epithelium, when grown intermingled with neonatal vaginal stroma, develops histotypic features of vagina. In contrast, vaginal epithelium, if grown with
P.1001
neonatal uterine stroma, develops a uterine phenotype. The final epithelium has cytosolic proteins characteristic of the induced rather than original epithelial source.
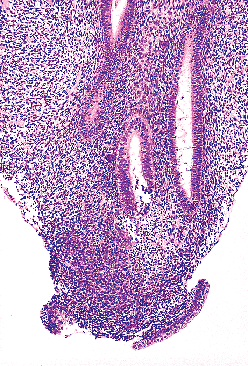 |
Figure 40.2 Single m llerian tube flanked by two wolffian ducts (circa day 67). The cytologic features of the cells comprising both types of duct are indistinguishable on light microscopy at early stages of development. Reprinted with permission from: Robboy SJ, Ellington KS. Pathology of the Female Genital Tract in Kodachrome Slides. 2nd ed. Durham, NC: Gyn-Path Associates; 1996. |
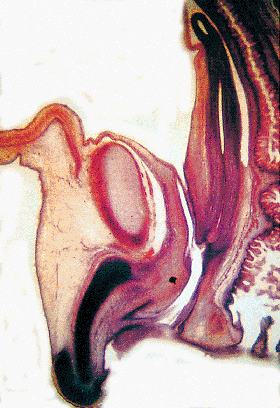 |
Figure 40.3 Sagittal section of pelvis at 16 weeks, showing vagina, bladder, urethra, and urogenital sinus. The lower genital tract is a straight longitudinal tube; and, at this stage, the vagina and cervix cannot be distinguished. Reprinted with permission from: Robboy SJ, Ellington KS. Pathology of the Female Genital Tract in Kodachrome Slides. 2nd ed. Durham, NC: Gyn-Path Associates; 1996. |
Additional evidence that the vaginal stroma plays a key role in epithelial differences comes from embryologic appearance of estrogen receptors. Among various body organs tested, the receptors are detected only in the genital tract. They appear in the mesenchyme at the tenth week, just before the m llerian epithelium is normally replaced by squamous epithelium of sinus origin (8). Epithelial labeling appears later (sixteenth week), at the time when the sinus (squamous) epithelium begins to mature and accumulate large quantities of glycogen. Interestingly, the stroma probably plays a key role in the action of MIS as well; the receptor for MIS is located in the stroma surrounding the m llerian duct rather than in the m llerian epithelium (11).
The developmental morphology of the vaginal mucosa and the inductive properties of the stroma supporting the vaginal mucosa are complex. For example, a hitherto unrecognized band of subepithelial stroma was described in 1973 that extended from the endocervix to the vulva (Figure 40.4) (12). It is 0.5 to 5 mm thick in mature females and most prominent in the endocervix. It is from this zone that fibroepithelial polyps seem to arise, an entity of no apparent physiologic function but which clinically should not be confused with malignant tumor (13,14).
Should the squamous epithelium described above fail during the critical weeks of embryonic life to replace the
P.1002
original columnar cells lining the vagina, the columnar cells remain in an arrested state of development until sometime around puberty, when they may further differentiate into the adult-type epithelium usually seen in biopsy material. It is conjectured that the tuboendometrial type of epithelium is the basic form of epithelium supported by the vaginal mesenchyme. In fact, it may be that this type of epithelium is the basic type of m llerian epithelium supported throughout the female genital tract, manifesting as serous cells in the uterine tube, endometrioid cells in the uterine corpus, and tuboendometrial cells in the vagina cells that are quite similar. In the cervix, a tuboendometrial layer of epithelium also lies deep to and as a cuff about the luminal layer of mucinous epithelium (15); the tuboendometrial layer, which is continuous with the lining of the corpus, is readily observed in hysterectomy specimens but is located too deep to be detected on biopsy. In fetuses where the vaginal lining has become squamous (older than 10 weeks), the inner stromal zone is obvious in the uterine tube, endometrium, and endocervix and tapers, appearing to end at the squamocolumnar junction of the cervix and vagina. Part of this layer may correspond to the most superficial stromal layer in the adult vagina described above. The original tuboendometrial layer is the origin of glandular remnants in the vagina of adults (adenosis).
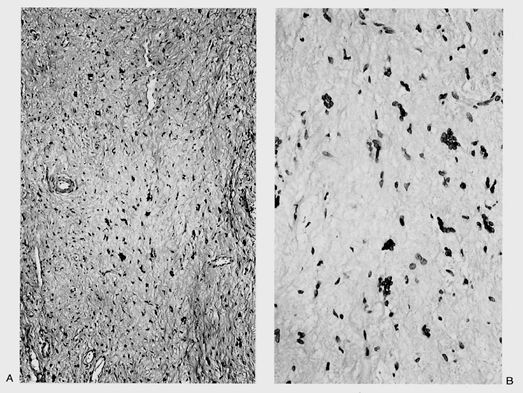 |
Figure 40.4 Bandlike area composed of loose connective tissue and multinucleated cells running the length of the vagina. A. Low-power view. B. High-power view. Reprinted with permission from: Fu YS. Pathology of the Uterine Cervix, Vagina and Vulva. 2nd ed. Philadelphia: WB Saunders; 2002. |
Gross Features
The vagina (from the Latin for sheath) extends from the vestibule of the vulva to the uterus, lying posterior (dorsal) to the urinary bladder and anterior (ventral) to the rectum. It's axis averages 30 degrees with the vertical, arching slightly posteriocranially, and more than 90 degrees with the uterus (Figure 40.5). The anterior wall averages 8 cm in length and the posterior wall 11 cm, with the cervix filling the 3 cm difference. In early life, the vagina is constricted distally, dilated in the middle, and narrowed proximally. It surrounds the exocervix and forms vaultlike fornices between its cervical attachment and the lateral wall. In the adult, the anterior and posterior walls are slack and remain in contact with each other, whereas the lateral walls remain fairly rigid and separated. This is thought to give an H-shaped appearance to the vaginal canal on cross section (16), although with three-dimensional imaging with magnetic resonance imaging (MRI), a W shape is now also recognized (1). During intercourse, the position of the uterus and bladder change relative to the vagina (17).
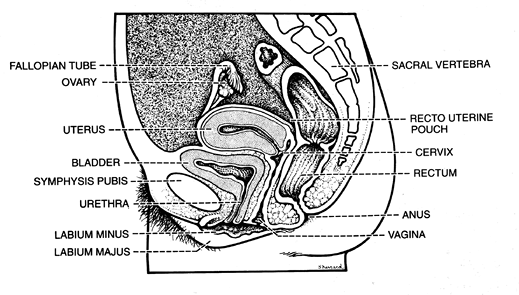 |
Figure 40.5 Structural relationships of the vagina. Its axis forms an angle of more than 90 degrees with the uterus. Reprinted with permission from: Zaino R, Robboy SJ, Kurman RJ. Diseases of the vagina. In: Kurman RJ, ed. Blaustein's Pathology of the Female Genital Tract. New York: Springer-Verlag; 2002:151 206. |
Posteriorly, the upper one-fourth of the vagina is related to the rectouterine space (i.e., the cul-de-sac or pouch of Douglas), which is covered with peritoneum. The middle half of the vagina is closely apposed to the rectum, separated only by fibrofatty adventitia and the rectovaginal septum. The lower one-fourth of the vagina is separated from the anal canal by anal and rectal sphincters, as well as the interposing perineal body, which contains the origin of the bulbocavernous and superficial transverse perineal muscles.
The urinary bladder and urethra lie anterior to the vagina. The urethra courses approximately one-third of its length on the vagina and then enters into the vaginal wall to become an inseparable part of it, usually terminating with its external meatus at the introitus. Typically, the meatus is outwardly directed, but not uncommonly it is directed into the outermost vagina. The ureters course along both sides of the upper one-third of the vagina until entering the bladder wall.
The vagina opens into the vestibule formed from the urogenital sinus. The vestibule lies beneath the urethra and between the inner margins of the labia minora. The vagina, urethra, and ducts of Bartholin's glands open into the vestibule. The size and shape of the vaginal orifice are related to the state of the hymen. When the inner edges of
P.1003
the hymen are apposed, the vaginal opening resembles a cleft. When stretched, the hymen may persist in the form of a ringlike structure about the readily recognized vaginal orifice. (See Chapter 39 for the anatomy of the hymenal region.)
Anatomy
Ligaments
The vaginal supports are intimately related to the uterus, urethra, bladder, and rectum (18). The lateral supports are called cardinal ligaments, the posterior supports sacrouterine ligaments. They originate where the isthmus of the uterine cervix and the uterine corpus meet and course outward, fanlike to the lateral and posterior pelvic walls. The isthmic fibers turn upward onto the uterus and downward onto the vagina. These ligaments, the connective tissues surrounding the vessels on the lateral vaginal walls, and the close proximity of the rectum, bladder, and urethra all contribute to support the vagina within the pelvis.
Blood Supply
The blood supply to the vagina is complex, with extensive anastomoses maintaining an adequate blood supply to all areas of the vagina in the event that injury restricts any single route of supply. The internal iliac (hypogastric) artery is the principal source of blood cranially as branches of the uterine arteries and caudally as branches of the middle hemorrhoidal arteries and pudendal arteries. Beginning cranially, the uterine artery gives off a descending branch, the cervicovaginal artery. Several branches supply the cervix. Lower branches supply the vagina. The vaginal arteries, which lie lateral to the vagina, send branches to both the anterior and posterior surfaces. The lower vagina receives its supply from ascending branches of the middle hemorrhoidal arteries and pudendal arteries, which also divide to send rami to the anterior and posterior vaginal walls. In toto, the extensive rami form a plexus around the vagina from which arise the median arteries, the azygos vaginal arteries on the anterior and posterior walls. A rich venous plexus also surrounds the vagina and communicates with the vesicle, pudendal, and hemorrhoidal venous plexuses, which empty into the internal iliac veins.
Nerves
The autonomic system of the pelvis originates in the superior hypogastric plexus. The middle hypogastric plexus, which passes into the pelvis, divides at the level of the S1 (sacral) vertebra into branches that pass to both sides of the pelvis and initiate the inferior hypogastric plexus. The inferior hypogastric plexus, a divided continuation of the middle hypogastric plexus, the superior hypogastric plexus, and the presacral nerve, descends into the pelvis in a position posterior to the common iliac artery and anterior to the sacral plexus; it curves laterally and finally enters the sacrouterine ligament. The medial segment of the primary division of the sacral nerves (S2 S5), as it sends fibers into the pelvic plexus located within the sacrouterine folds, appears to contain both sympathetic (inferior hypogastric plexus) and parasympathetic (nervi erigentes) components. An extension of this plexus, located in the base of the broad ligament and supplied by the middle vesical artery, contains many ganglia. Most nerves enter the uterus near the isthmus. A lesser number descend along the lateral vagina, a pattern similar to the arteries that supply the vagina.
Lymphatic Drainage
The vaginal lymphatic system, despite the simplified view given here, is highly variable (Figure 40.6) (19). The lymphatics begin as a delicate plexus of small channels involving the entire mucosa and lamina propria and then drain into a deep muscular network. They terminate in a perivaginal plexus from which arise collecting trunks, which themselves coalesce into several larger channels.
The lymph drainage follows patterns that reflect functionally diverse geographic regions. The lymphatics of the upper anterior wall join those of the cervix, where they follow the cervical vessels to the uterine artery and accompany it to terminate in the medial chain of the external iliac
P.1004
nodes. The lymph from the posterior vagina drains into deep pelvic, rectal, and aortic nodes. The lymphatics of the lower vagina, which also include the hymenal region, follow two distinct courses. One passes to the interiliac nodes in company with the upper vaginal discharge. The other traverses the paravesical space, carrying lymph to the deepest portions of the pelvis and draining into the inferior gluteal nodes near the origin of the vaginal or internal pudendal artery. The channels that anastomose with those of the vulva drain to the superficial iliac nodes. In summary, as a practical matter, one can generalize that tumors in the upper vagina will spread like cervical carcinoma to involve obturator and both internal and external iliac nodes. In contrast, tumors in the lower vagina will tend to involve superficial iliac (inguinal) and deep pelvic nodes, similar to vulvar cancer.
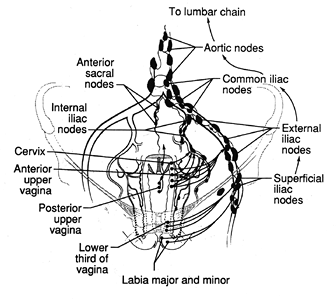 |
Figure 40.6 Lymphatic drainage of the vagina. Reprinted with permission from: Moore TR, Reiter RC, Rebar RW, Baker VV, eds. Gynecology and Obstetrics: A Longitudinal Approach. New York: Churchill Livingstone; 1993. |
Light Microscopy
Epithelium
The vaginal wall consists of three principal layers: mucosa (epithelial and submucosal stroma), muscle, and adventitia. The epithelium is about 0.4 mm thick and, on gross examination, exhibits a characteristic pattern of folds or rugae separated by furrows of variable depth. There are two longitudinal (anterior and posterior) and multiple transverse furrows. The rugal pattern of the vaginal mucosa, which contributes to the organ's elasticity, produces an undulating appearance on microscopic examination in contrast to the flat surface of the cervix. The rugae, which are more prominent in nulliparous than multiparous women, reinforce the gripping effect of the levator ani and vaginal constrictor muscles during intercourse. Nonkeratinized glycogenated squamous epithelium lines the luminal surface in a manner similar to cervical epithelium. The normal vaginal mucosa lacks glands. Its surface is lubricated both by fluids that pass directly through the mucosa and by cervical mucus.
The mature, stratified squamous epithelium can be subdivided into several layers, typical of squamous epithelia elsewhere in the body (Figure 40.7). From the base to the surface, they are the deep, intermediate, and superficial zones. The deep zone contains the basal cell layer and, above this, the parabasal layer. Both are the active proliferative compartments or germinal beds, as shown by the Ki-67 antigen, which is demonstrable during late G1, G2, and M phases of the cell cycle (Figure 40.8). The basal cell layer consists of a single layer of columnar-like cells, approximately 10 m thick, the long axis of which is vertically arranged. The cells have a basophilic cytoplasm and relatively large oval nuclei. Mitoses may be present. Occasional melanocytes also are found.
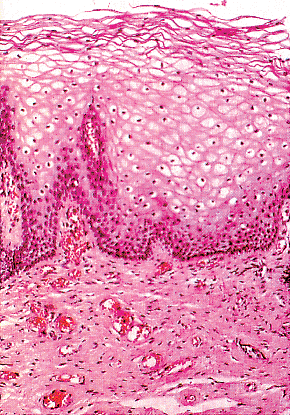 |
Figure 40.7 Mucosa of the adult vagina. Mature cells with glycogenic cytoplasm and pyknotic nuclei occupy most of the epithelial thickness. There is a single layer of dark basal cells and three to four layers of intermediate cells. Reprinted with permission from: Robboy SJ, Ellington KS. Pathology of the Female Genital Tract in Kodachrome Slides. 2nd ed. Durham, NC: Gyn-Path Associates; 1996. |
The parabasal layer is poorly demarcated from the overlying cell layers. It consists usually of about two layers of small polygonal cells, having a total 14 m thickness, often with intercellular bridges. The cells have basophilic cytoplasm, a relatively large, centrally placed, round nucleus, and occasional mitoses.
The intermediate cell layer is of variable thickness. The cells have prominent intercellular bridges, a naviculate configuration, and a long cell axis paralleling the surface. The cytoplasm is basophilic, although some glycogen may be present. The nuclei are round, oval, or irregular, with finely granular chromatin. This layer of cells has about 10 rows of cells of about 100 m thickness.
The superficial layer is also of variable thickness. The cells are polygonal when viewed from above and flattened when viewed in cross section. The cytoplasm is acidophilic, and the nuclei are centrally located, small, round, and pyknotic. Keratohyalin granules are sometimes seen in the cytoplasm. This layer also contains about 10 rows of squamous cells.
Relatively little is known about the normal components of the epithelium itself. The submucosa contains a variety of mononuclear cells demonstrable by immunocytochemical
P.1005
methods. The dendritic processes of Langerhans cells (about 4 per high power field) are distributed throughout the mucosa (20). We have found them largely in the deeper layers, but they can extend into the superficial fields (Figure 40.9). Both T8 and, to a lesser degree, T4 lymphocytes are also frequently found, whereas macrophages and B lymphocytes are relatively uncommon.
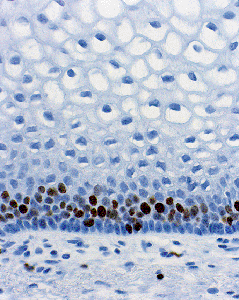 |
Figure 40.8 Ki-67 antigen, demonstrable during late G1, G2, and M phases of the cell cycle, in the basal and parabasal layers of the normal vaginal mucosa. Reprinted with permission from: Robboy SJ, Ellington KS. Pathology of the Female Genital Tract in Kodachrome Slides. 2nd ed. Durham, NC: Gyn-Path Associates; 1996. |
Epithelial Response to Hormones
Vaginal epithelial cells proliferate and mature in response to stimulation by ovarian or exogenous estrogenic hormones. Hence, it would seem that the total number of squamous cell layers would vary greatly during the normal menstrual cycle and as a woman passes through the various stages of the life cycle, that is, birth, childhood, reproduction, and the postmenopausal years. Earlier work indicated the high occurred at ovulation (average 45 layers). It built slowly during the proliferative phase and averaged 22 on day 10. After ovulation, the number receded to 33 on day 19 and to 23 on day 24 (21,22). More recent workers found there was a statistically significant decrease from 27.8 on days 1 to 5 and 28.1 on days 7 to 12 to 26.0 on days 19 to 24, but clinically this appears to be of little import (20).
Without hormonal stimulation, the cells atrophy (Figure 40.10). At the peak of estrogenic activity (i.e., just before ovulation), the superficial cells with abundant intracytoplasmic glycogen predominate, both on histologic section and in smears (Figure 40.11). Lactobacilli metabolize the glycogen normally present in the vagina to lactic acid, which maintains an acid vaginal pH (about pH 4.4 during the late proliferative and secretory phases) (23).
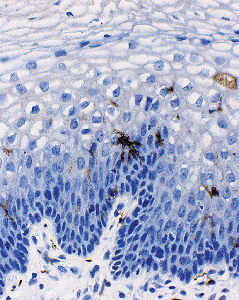 |
Figure 40.9 The dendritic processes of Langerhans cells. Reprinted with permission from: Robboy SJ, Ellington KS. Pathology of the Female Genital Tract in Kodachrome Slides. 2nd ed. Durham, NC: Gyn-Path Associates; 1996. |
Progesterone inhibits maturation of the vaginal epithelium. Consequently, intermediate cells predominate when the circulating levels of progesterone are high; for example, during the postovulatory phase of the menstrual cycle or during pregnancy. Estrogenic activity is low or absent before puberty and after the menopause; the vaginal
P.1006
epithelium fails to mature and hence remains thin. Parabasal and intermediate cells predominate in the vaginal smear. In the newborn child, the vaginal epithelium is frequently mature because of the influence of placental estrogens (Figure 40.12). Quantitative studies measuring the rate of change in the maturation index in the infant's vagina from birth to the atrophic state indicate that vaginal cells replace themselves in less than two weeks; that is, the time required for basal cells to work their way up and become desquamated superficial cells. Studies of the exocervix indicate that turnover there is also rapid (24).
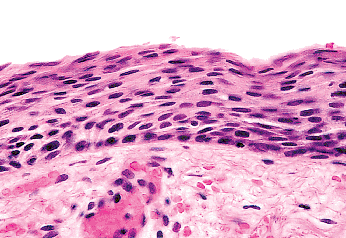 |
Figure 40.10 Atrophic vagina. Reprinted with permission from: Robboy SJ, Ellington KS. Pathology of the Female Genital Tract in Kodachrome Slides. 2nd ed. Durham, NC: Gyn-Path Associates; 1996. |
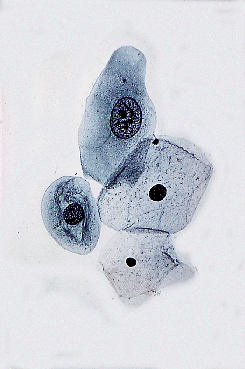 |
Figure 40.11 Vaginal smear, showing basal cell, two intermediate cells, and a superficial cell (Papanicolaou stain). Reprinted with permission from: Robboy SJ, Ellington KS. Pathology of the Female Genital Tract in Kodachrome Slides. 2nd ed. Durham, NC: Gyn-Path Associates; 1996. |
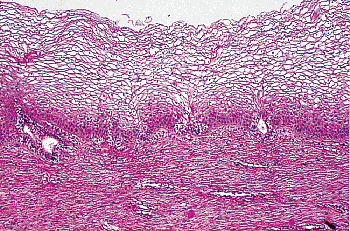 |
Figure 40.12 Vaginal mucosa of a near-term fetus. Mature cells predominate and cannot be distinguished from that of the adult (compare with Figure 40.7). In addition, two adenotic glands of the embryonic type are present. Reprinted with permission from: Robboy SJ, Ellington KS. Pathology of the Female Genital Tract in Kodachrome Slides. 2nd ed. Durham, NC: Gyn-Path Associates; 1996. |
The submucosa, or lamina propria, lies beneath the squamous epithelium. It contains elastic fibers and a rich venous and lymphatic network. Sometimes the superficial lamina propria discloses a bandlike zone of loose connective tissue that contains atypical polygonal to stellate stromal cells with scant cytoplasm (Figure 40.4). Many cells are multinucleated or have multilobulated hyperchromatic nuclei. Few are mononucleate. Mitoses are not observed. These atypical stromal cells are thought to give rise to fibroepithelial polyps, which have been observed within the cervix, vagina, and vulva. They have been shown to be fibroblastic in origin.
Vaginal Wall and Adventitia
The vaginal musculature is continuous with that of the uterus. The outer layers in muscle of both the uterus and vagina run longitudinally to pass out onto the lateral pelvic wall to form the superior and inferior surfaces, respectively, of the cardinal ligaments. The longitudinal muscle fibers continue to course the length of the vagina to the region of the hymenal ring, where they gradually disappear in the connective tissue. On the anterior vaginal wall, the longitudinal muscle fibers are displaced by the urethra more than diminished in number. The inner muscle layer of the vagina forms a spiral-like course, appearing in microscopic sections as somewhat circular in direction.
The adventitia is a thin coat of dense connective tissue adjoining the muscularis. The connective tissue of the adventitia merges with the stroma, connecting the vagina to the adjacent structures. This layer contains the many veins, lymphatics, nerve bundles, and small groups of nerve cells.
Ultrastructure
On ultrastructural examination, the component layers are not sharply demarcated from each other. Rather, they may be somewhat difficult to distinguish because each layer has ill-defined limits and displays gradual changes in structure (25). In general the ultrastructural changes observed by transmission electron microscopy resemble those of the exocervix and mucosal squamous cells and are not further covered herein.
On scanning electron microscopic examination, the superficial cells appear large (50 m in greatest dimension) and polygonal. The intercellular edges are narrow and dense and protrude slightly. The pattern of fine webbing and anastomotic intercellular bridges typifies nonkeratinized
P.1007
squamous epithelium, such as that observed in buccal mucosa. The key structure on the cell surface is the microridge, or in reality myriad microridges, which are interanastomotic longitudinal elevations of the plasma membrane 0.2 nm long and 0.1 nm high (Figure 40.13). Arranged in dense convolutions, they tie one cell to another, operating in a zipper-fastener principle. They are thought to provide surface adhesion. Desmosomes are prominent in these areas.
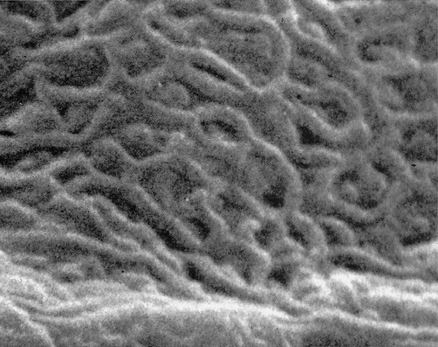 |
Figure 40.13 Intricate network of microridges on surface plasma membrane of most superficial squamous cells living in the normal vaginal lumen. Scanning electron microscopy (original magnification 10,000). Reprinted with permission from: Ferenczy A, Richart RM. Female Reproductive System: Dynamics of Scan and Transmission Electron Microscopy. New York: John Wiley; 1974. |
Microridge formation depends on the topographic configuration of disulfide-rich keratin or keratin precursors, which are absent in immature precursor cells, such as intermediate cells and young metaplastic squamous cells. From midcycle and early in the luteal phase, intercellular grooves widen. Porelike widening (porosites) of the intercellular crevices takes place where several cells interconnect. This porosity is thought to enhance continuity of the intercellular space system of the vaginal epithelium and the vaginal surface, thus permitting free passage of vaginal lubricating fluid.
Information about changes at the biochemical and immunological levels is relatively little, but clearly the cells at various levels show dramatic differences. Part may reflect degrees of cellular maturity. Part may reflect subcellular specialization. Like other epithelia, both squamous and glandular alike, the pattern of cytokeratin expression reflects the state and nature of differentiation of the epithelial cells. The vaginal mucosa reflects that of other nonkeratinized squamous epithelium. Cytokeratin 14 (CK14) is demonstrable in the basal layer and CK13 in the parabasal layer, the latter supporting a subpopulation of cells that express CK10 (26). In ways uncertain, the differentiating cells show changes that have other effects. One is its influence on bacterial binding, such that cellular receptors that develop in the differentiated cells enhance adhesion of pathogenic bacteria. As one example, differentiated vaginal mucosal cells express receptors to Escherichia coli type 1 pili, which are surface-adhesive organelles (26). Such colonization where fecal E. coli are present in the vaginal introitus may be a key initial event leading to acute urinary infection. Another example is the presence of a surfactant protein (SP-A), which the human vaginal epithelial cells secrete. This factor, an important host defense, acts to facilitate micoorganism phagocytosis (27,28).
Differential Diagnosis and Special Anatomy
Wolffian Ducts
The wolffian duct, known otherwise as the mesonephric duct or Gartner's duct, is vestigial in the adult female (Figure 40.14). It begins to irreversibly wither if not stimulated to develop by testosterone before the thirteenth week postconception. This paired duct is most commonly situated in the lateral vaginal walls, although we have encountered it in all areas. Where encountered by chance in a radical vaginectomy specimen, the ducts are virtually always invisible grossly. Mitoses are absent. Usually it is a small duct or clusters of small glands about a duct. The lumen is filled frequently with a deeply eosinophilic, hyalinized secretion. The single layer of cells lining the duct is primarily composed of the cell nucleus. The cytoplasm is scant, relatively translucent, and lacks cilia. The nuclei frequently overlap. The chromatin is strikingly bland. On a
P.1008
clinical basis, individual ducts occasionally become cystic and macroscopically visible. In the cervix, these ducts rarely appear diffusely throughout the wall and appear as mesonephric hyperplasia or even adenoma (29). We also have seen rare cases where the neoplasm present appeared to be a true wolffian duct carcinoma.
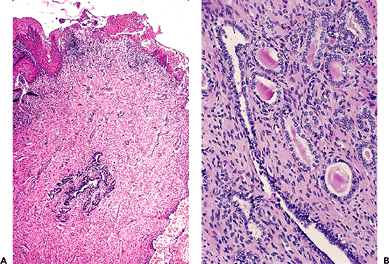 |
Figure 40.14 A. Vestigial wolffian duct remnants, deep in wall. B. Detail of central duct and arborized ductal terminals with eosinophilic secretions. Reprinted with permission from: Robboy SJ, Ellington KS. Pathology of the Female Genital Tract in Kodachrome Slides. 2nd ed. Durham, NC: Gyn-Path Associates; 1996. |
Remnants of M llerian Duct Epithelium (Adenosis)
The DES story began in 1938 when this nonsteroidal estrogen was synthesized and then gained popularity for the treatment of high-risk pregnancy. By 1971, up to two million women had taken the drug, at which time it was linked to the extremely rare development of clear cell adenocarcinoma of the vagina in young female offspring. Subsequently, about one-third of the exposed young women were found to have adenosis (presence of glandular tissue in the vagina). Both retrospective and prospective studies have shown that adenosis can be found in nonexposed women also, albeit rarely. In both exposed and nonexposed women, the adenosis is related to embryonic m llerian tissue that has remained entrapped and not been replaced by squamous epithelium during fetal life.
Adenosis appears in three forms. One type, the embryonic form, is exceedingly rare. The other two are the adult and common forms. In adenosis found during fetal life and in stillborns, but only rarely in adults, the glands are embryonic in character (Figures 40.12, 40.15) (30). They are small, usually at the epithelial stromal interface, and are characterized by individual cells with small basal nuclei and copious bland cytoplasm that does not stain with either periodic acid-Schiff or mucicarmine.
It is believed that adenosis takes on its adult forms in women some time during puberty (31,32). Mucinous columnar cells, which by light and electron microscopy resemble those of the normal endocervical mucosa, comprise the glandular epithelium most frequently encountered as adenosis (62% of biopsy specimens with vaginal adenosis). This epithelium, because it frequently lines the surface of the vagina, is the type most commonly observed by colposcopy. Commonly, the mucinous columnar cells also line glands embedded in the lamina propria. This form of epithelium gives rise to the progestin-stimulated lesion, microglandular hyperplasia of the vagina (33).
Dark cells and light cells, often ciliated and resembling the lining cells of the uterine tube and endometrium, are found in 21% of specimens in the upper vagina with adenosis. This form of adenosis has been called tuboendometrial, although serous might be equally appropriate. The cells are usually found in glands in the lamina propria and not on the vaginal surface. Although adenosis
P.1009
in the lower vagina is rare in absolute number, the percentage of biopsy specimens with adenosis that exhibits tuboendometrial rather than mucinous cells increases markedly in frequency in comparison with the more cranial aspects of the vagina. The tuboendometrial cell, which is benign, is the cell that we believe is related to clear cell adenocarcinoma, possibly through atypical adenosis, a transitional form (15,34). Mucinous glands and mucinous pools or droplets are encountered frequently in the same biopsy specimen; mucinous and tuboendometrial cells are found together only occasionally in biopsy material. The tuboendometrial form of adenosis is the instrumental type of glandular cell induced by all regions of the m llerian duct, be it uterine tube, uterus, or vagina, whereas the mucinous cell is generally specific to the endocervix or, after DES exposure, to the deformed region of the cervix, which becomes ill-defined and includes what appears to be the upper vagina.
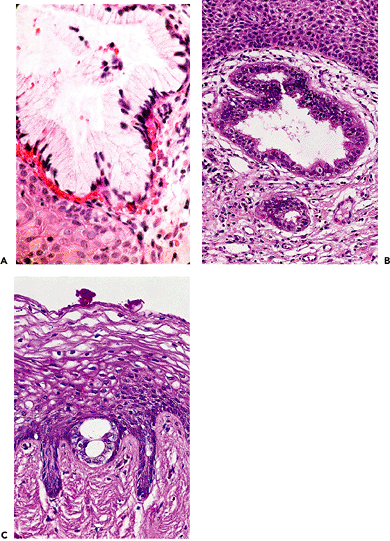 |
Figure 40.15 Vaginal adenosis in which the glandular epithelium is mucinous (A), tuboendometrial (B), or of the immature embryonic type (C). Reprinted with permission from: Robboy SJ, Ellington KS. Pathology of the Female Genital Tract in Kodachrome Slides. 2nd ed. Durham, NC: Gyn-Path Associates; 1996. |
P.1010
References
1. Barnhart KT, Pretorius ES, Malamud D. Lesson learned and dispelled myths: three-dimensional imaging of the human vagina. Fertil Steril 2004;81:1383 1384.
2. Robboy SJ, Bentley RC, Russell P, Anderson MC. Embryology and Disorders of Sexual Development. In: Robboy SJ, Anderson MC, Russell P, eds. Pathology of the Female Reproductive Tract. London: Churchill Livingstone; 2002:819 860.
3. Aittomaki K, Eroila H, Kajanoja P. A population-based study of the incidence of mullerian aplasia in Finland. Fertil Steril 2001;76:624 625.
4. Lawrence WD, Whitaker D, Sugimura H, Cunha GR, Dickersin GR, Robboy SJ. An ultrastructural study of the developing urogenital tract in early human fetuses. Am J Obstet Gynecol 1992;167:185 193.
5. MacLaughlin DT, Donahoe PK. Mechanisms of disease: Sex determination and differentiation. N Eng J Med 2004;350:367 378.
6. Taguchi O, Cunha GR, Lawrence WD, Robboy SJ. Timing and irreversibility of Mullerian duct inhibition in the embryonic reproductive tract of the human male. Dev Biol 1984;106:394 398.
7. Ulfelder H, Robboy SJ. The embryologic development of the human vagina. Am J Obstet Gynecol 1976;126:769 776.
8. Taguchi O, Cunha GR, Robboy SJ. Expression of nuclear estrogen-binding sites within developing human fetal vagina and urogenital sinus. Am J Anat 1986;177:473 480.
9. Terruhn V. A study of impression moulds of the genital tract of female fetuses. Arch Gynecol 1980;229:207 217.
10. Kurita T, Cooke PS, Cunha GR. Epithelial-stromal tissue interaction in paramesonephric (Mullerian) epithelial differentiation. Dev Biol 2001;240:194 211.
11. Tsuji M, Shima H, Yonemura CY, Brody J, Donahoe PK, Cunha GR. Effect of human recombinant mullerian inhibiting substance on isolated epithelial and mesenchymal cells during mullerian duct regression in the rat. Endocrinology 1992;131:1481 1488.
12. Elliott GB, Elliott JD. Superficial stromal reactions of lower genital tract. Arch Pathol 1973;95:100 101.
13. Abdul-Karim FW, Cohen RE. Atypical stromal cells of lower female genital tract. Histopathology 1990;17:249 253.
14. Robboy SJ, Anderson MC, Russell P. Vagina. In: Robboy SJ, Anderson MC, Russell P, eds. Pathology of the Female Reproductive Tract. London: Churchill Livingstone; 2002:75 104.
15. Robboy SJ, Welch WR, Young RH, Truslow GY, Herbst AL, Scully RE. Topographic relation of cervical ectropion and vaginal adenosis to clear cell adenocarcinoma. Obstet Gynecol 1982;60:546 551.
16. Nichols DH, Randall CL. Pelvic anatomy of the living. In: Nichols DH, Randall CL, eds. Vaginal Surgery. 4th ed. Baltimore: Williams & Wilkins; 1996:1 42.
17. Faix A, Lapray JF, Courtieu C, Maubon A, Lanfrey K. Magnetic resonance imaging of sexual intercourse: initial experience. J Sex Marital Ther 2001;27:475 482.
18. Mostwin JL. Current concepts of female pelvic anatomy and physiology. Urol Clin North Am 1991;18:175 195.
19. Moore TR, Reiter RC, Rebar RW, Baker VV, eds. Gynecology and Obstetrics: A Longitudinal Approach. London: Churchill Livingstone; 1993.
20. Patton DL, Thwin SS, Meier A, Hooton TM, Stapleton AE, Eschenbach DA. Epithelial cell layer thickness and immune cell populations in the normal human vagina at different stages of the menstrual cycle. Am J Obstet Gynecol 2000;183:967 973.
21. Burgos MH, Roig de Vargas-Linares CE. Ultrastructure of the vaginal mucosa. In: Hafez ESE, Evans TN, eds. The Human Vagina. New York: North-Holland; 1978:63 93.
22. Steger RW, Hafez ESE. Age associated changes in vagina. In: Hafez ESE, Evans TN, eds. The Human Vagina. New York: North-Holland; 1978:95 108.
23. Eschenbach DA, Thwin SS, Patton DL, et al. Influence of the normal menstrual cycle on vaginal tissue, discharge, and microflora. Clin Infect Dis 2000;30:901 907.
24. Linhartova A. The height and structure of the cervical squamous epithelium in foetuses, newborns, and girls. Cervix Low Female Genital Tract 1989;7:37 48.
25. Ferenczy A, Richart RM. Female Reproductive System: Dynamics of Scan and Transmission Electron Microscopy. New York: John Wiley; 1974.
26. Klumpp DJ, Forrestal SG, Karr JE, Mudge CS, Anderson BE, Schaeffer AJ. Epithelial differentiation promotes the adherence of type 1-piliated Escherichia coli to human vaginal cells. J Infect Dis 2002;186:1631 1638.
27. MacNeill C, Umstead TM, Phelps DS, et al. Surfactant protein A, an innate immune factor, is expressed in the vaginal mucosa and is present in vaginal lavage fluid. Immunology 2004;111:91 99.
28. Wira CR, Fahey JV. The innate immune system: gatekeeper to the female reproductive tract. Immunology 2004;111:13 15.
29. Ferry JA, Scully RE. Mesonephric remnants, hyperplasia, and neoplasia in the uterine cervix: a study of 49 cases. Am J Surg Pathol 1990;14:1100 1111.
30. Robboy SJ, Hill EC, Sandberg EC, Czernobilsky B. Vaginal adenosis in women born prior to the diethylstilbestrol era. Hum Pathol 1986;17:488 492.
31. Robboy SJ. A hypothetic mechanism of diethylstilbestrol(DES)-induced anomalies in exposed progeny. Hum Pathol 1983;14:831 833.
32. Robboy SJ, Kaufman RH, Prat J, et al. Pathologic findings in young women enrolled in the National Cooperative Diethylstilbestrol Adenosis (DESAD) project. Obstet Gynecol 1979;53:309 317.
33. Robboy SJ, Welch WR. Microglandular hyperplasia in vaginal adenosis associated with oral contraceptives and prenatal diethylstilbestrol exposure. Obstet Gynecol 1977;49:430 434.
34. Robboy SJ, Young RH, Welch WR, et al. Atypical vaginal adenosis and cervical ectropion. Association with clear cell adenocarcinoma in diethylstilbestrol-exposed offspring. Cancer 1984;54:869 875.
- Integration Strategies and Tactics for Information Technology Governance
- Assessing Business-IT Alignment Maturity
- A View on Knowledge Management: Utilizing a Balanced Scorecard Methodology for Analyzing Knowledge Metrics
- Measuring ROI in E-Commerce Applications: Analysis to Action
- Technical Issues Related to IT Governance Tactics: Product Metrics, Measurements and Process Control