25 - Colon
Editors: Mills, Stacey E.
Title: Histology for Pathologists, 3rd Edition
Copyright 2007 Lippincott Williams & Wilkins
> Table of Contents > VIII - Hematopoietic System > 32 - Spleen
32
Spleen
J. Han J. M. van Krieken
Attilio Orazi
Introduction
Historically, the human spleen has attracted attention from poets as a producer of melancholy. Galen (131 201 AD) called the human spleen an enigmatic organ, a notion that has persisted for a long time. In the seventeenth century, Malpighi described, macroscopically, the splenic lymphoid follicles as white pulp against a background of red pulp. In 1857, Billroth published one of the first histologic studies of the human spleen in which he divided the red pulp into cord tissue and venous sinuses (1). Still, until the second half of the twentieth century, the spleen was considered a rather useless reservoir for blood cells and was hardly studied. In the 1970s, by using electron microscopy, Weiss was able to elucidate the fine structure of the organ, which gave insights into the red pulp function (2,3). During the same period, Nieuwenhuis, Ford, and Keuning performed immunologic function studies on rat spleen (4,5,6), and Veerman published a detailed description of the white pulp of the rat spleen (7). A complete summary of the organization and functions of the spleen is provided by Weiss in 1988, in the sixth and last edition of his Textbook of Histology (Cell and Tissue Biology) (8).
Nevertheless, many pathologists still lack a clear understanding of the normal histology and functions of the human spleen. This is due to several reasons. The organ is extremely vulnerable to autolysis, which often makes histologic findings in postmortem specimens difficult to interpret and of limited teaching value. Surgically removed spleens are suitable, if processed without delay. However, since the number of splenectomies performed in most institutions is relatively scarce, it is not surprising that pathologists may feel uncomfortable when interpretating splenic pathology as a result of a lack of familiarity with splenic histologic features.
Another source of confusion with respect to the structure and function of the human spleen is in the terminology and definitions applied to this organ, which are still largely based on studies of animal spleens. The human and
P.784
animal spleens do not have an identical architecture; for example, in the human spleen, the periarteriolar lymphocyte sheath and the marginal sinus as described in rodent spleens are not present. Furthermore, certain definitions (e.g., of the marginal zone) vary widely from author to author (9,10).
The next problem is the large variations that occur in the normal spleen. The spleen is a compartimentalized organ (Table 32.1). Stimulation of one of the many functions of the spleen can lead to morphologic changes in the compartment that is mainly responsible for that function. The normal spleen therefore can show wide variation. As one of us has shown in a previously published study, it is essential to define a normal control population if one undertakes histologic studies in the spleen in specific disorders (11). For example, a morphometric analysis showed that spleens removed incidentally during abdominal surgery (i.e., for highly selective vagotomy or early gastric cancer) differed from traumatically ruptured spleens; we therefore excluded the latter from our normal group. All these problems sometimes make it difficult to differentiate physiologic from pathologic changes.
Part of this chapter is based on a previous study, performed by one of the authors, of methylmethacrylate sections of more than 400 surgically removed human spleens and our immunohistochemical experience using frozen (12,13) and/or paraffin-embedded tissue.
Table 32.1 Summary of Splenic Histology, Function, and Relationship to Lymph Node Compartments | ||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||
Prenatal and Developmental Changes
During embryogenesis, the spleen can be recognized from about the fifth week of gestation, and blood vessels appear in it by the ninth week. Red and white pulp cannot be distinguished until the ninth month. The functional role of the spleen during prenatal development varies widely from that of the adult spleen, and this is reflected in the microscopic anatomy of the organ. Hematopoiesis was considered to take place in the fetal spleen (and liver) and to contribute largely to blood cell formation in the fetus until the sixth month of gestation, but it has been shown that, in fact, the spleen functions at most as a site of maturation for hematopoietic precursors derived from the peripheral blood (14,15). In adults, one may see foci of hematopoietic cells (extramedullary hematopoiesis) in the spleen in many reactive conditions (e.g., sepsis), as well as in disorders of the bone marrow associated with myelofibrosis. Extramedulary hematopoiesis as seen in the spleen is also referred to as myeloid metaplasia.
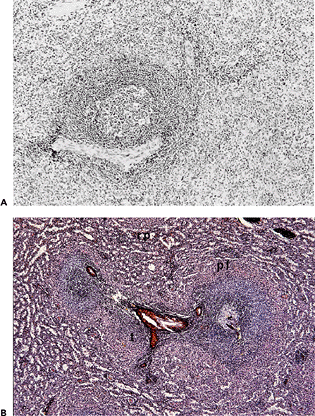 |
Figure 32.1 Spleen removed in idiopathic thrombocytopenic purpura. A. Formalin-fixed paraffin embedding (H&E, original magnification 40). B. Methylmethacrylate embedding (methenamine-silver/H&E, original magnification 40). Overview of red and white pulp showing central arteriole with T-cell area, a primary follicle, and a secondary follicle containing a germinal center. Note the absence of the marginal zone around the T-cell area and the presence of the erythrocyte-rich (pink) perifollicular zone surrounding both the T- and B-cell compartment of the white pulp. Note the lack of detail on the structure of the red pulp and the difficult discernable perifollicular zone in standard H&E section. (rp, red pulp; pf, perifollicular zone) |
P.785
The immune system develops during fetal growth, and this development continues after birth (16). This functional maturation is reflected by the morphology: until birth the splenic white pulp does not contain follicles and marginal zones. There are immature B cells in clusters and T cells scattered throughout the organ. Their numbers increase with the developmental age of the fetus; and, from the end of the second semester onward, B- and T-cell areas can be recognized (17,18). Phagocytosis can be demonstrated at the twelfth week of gestation (14,15).
Developmental changes of the spleen are very familiar. The presence of accessory spleens (so-called splenculi, small extra pieces of spleen tissue with the complete and normal histology of the red and white pulp) can be found in at least 25% of autopsies. In disorders being treated with splenectomy, these splenculi may lead to recurrence of the disease.
Rare but well known is the polysplenia associated with immotile cilia syndrome (19). In this syndrome, left-right orientation of thoracal and abdominal organs may be abnormal, and the spleen at the right side is often divided into many small pieces, generally having normal function. This is not to be confused with acquired splenosis, in which many small fragments of spleen are present after trauma. Congenital asplenia, which is exceedingly rare, is associated with abnormalities of the cardiovascular system.
Apoptosis
In the development of the spleen, apoptosis does not seem to play an important role, but the lymphoid compartment, as in other lymphoid tissues, shows extensive apoptosis, especially in the germinal centers of the B-cell follicles. This is illustrated in Figure 32.1, where the starry sky phenomenon can be observed. The starry sky cells are macrophages that phagocytose remnants of lymphocytes that are dying through apoptosis, generally because they
P.786
have an unsuccessful gene rearrangement of the antigen receptor or because of the fact that the produced immunoglobulin recognizes autoantigen. This physiologic process is important in the protection against autoimmune diseases. The Bcl-2 protein that protects against certain forms of apoptosis (see Chapter 1), which is expressed in most B and T cells, is lacking in germinal center B cells, rendering them susceptible for apoptosis. In follicular lymphoma the t(14;18) translocation leads to aberrant expression of Bcl-2 in the tumor cells. This is sometimes helpful in the recognition of follicular lymphoma in the spleen for the following reasons. Because the spleens of patients over about 20 years of age only rarely contain active germinal centers, the distinction between a primary follicle and follicular lymphoma can be difficult. Furthermore, the involvement of the spleen by follicular lymphoma is often nodular but does not lead to the disturbance of the architecture that is so noticeable in the lymph nodes involved by follicular lymphoma. Because spleens are often received after fixation, immunoglobulin light chain restriction is not possible by immunohistochemistry, immunofluorescence, or flow cytometry.
Apoptosis also plays an important role in mantaining a normal number and function of T cells. In cases of autoimmune lymphoproliferative syndrome (a pediatric disorder due to a genetic defect of FAS or FAS ligand that is associated with splenomegaly and autoimmunity), a decreased rate of apoptosis in T lymphocytes is responsible for the marked degree of lymphoid hyperplasia seen in the T-cell rich areas of the spleen.
Gross Features/Organ Weight
The human spleen is a bean-shaped organ surrounded by a smooth capsule covered by the peritoneum. In contrast to several species, the capsule does not contain smooth muscle fibers and therefore does not have the capability of undergoing contraction in response to acute blood loss. The spleen in animals such as dogs and cats has an important red blood cell reservoir function. By undergoing rapid contraction, the spleen can squeeze out its red blood cell reservoir and, by doing so, produce a rapid increase in the amount of circulating blood. The surface may be covered with fibrotic or even calcified plaques, the cause of which is unknown. It is not uncommon to find several grooves at the outer surface that have no clinical significance. The weight of the spleen is highly variable (20). In adults, the spleen generally weighs 150 to 250 g; but, in the elderly, the spleen is often substantially smaller, even when there is no apparent hypofunction.
On the cut surface, the red and white pulp can be discerned, the latter consisting of small (less than or equal to 2-mm) nodules. It is important to realize that involvement of the spleen in malignant lymphoma often is observed foremost in the white pulp (21), which becomes enlarged but often not to a great extent. Therefore, the spleen should be cut up into small sections (less than or equal to 5 mm).
Anatomy
Blood Supply
Blood reaches the spleen via the splenic artery, a large branch of the celiac artery, and enters the spleen through four to six branches; their number and location is, however, highly variable. Venous outflow occurs via four to six venous branches. These combine, within the lienorenal ligament to form the splenic vein, which drains into the portal vein (22). This is why portal hypertension can produce congestive splenomegaly. The blood flow within the spleen is highly specialized and relates to the different functions of the spleen.
Nerves
The spleen is innervated by nonmyelinated fibers from the major splanchnic nerves and the celiac plexus (23). These nerve fibers run along the splenic artery. Innervation in human spleens is less extensive than in cat and dog spleens, and this might be related to the important reservoir function of the spleen in these animals, as previously mentioned.
Lymphatics
No afferent lymphatic vessels are present in the spleen. Its lymph drainage occurs via hilar lymph nodes and lymph nodes in the gastrosplenic ligament. The lymph then flows through lymphatics along the splenic artery to the celiac lymph nodes along the celiac artery. The lymphatics in the spleen are described below.
Light Microscopy
Vascular Tree
After entering at the hilus, the splenic artery branches like a tree (24). Within the splenic parenchyma, these arterial branches, called trabecular arteries, are accompanied by veins and lymph vessels and surrounded by collagenous fibers. These vessels containing fibrous structures are usually referred to as trabeculae or septa, a term which is inappropriate to describe what in essence are perivascular collagen cuffs. Real, albeit short, true septa are also present in the spleen. These are connected to the capsule, lack inside vessels, and only extend for a short length into the splenic tissue. Foci of condensed reticular fibers devoid of vessels are found throughout the red pulp. The condensed reticulum
P.787
appears to be in direct continuity with the reticular meshwork of the surrounding red pulp; they may represent areas of collapse or involution of the red pulp tissue.
Trabecular arteries branch to form central arteries and arterioles that are no longer accompanied by veins and are surrounded not by a collagenous cuff but rather by lymphatic tissue predominantly composed of T lymphocytes. This lymphatic compartment, which is usually referred to as periarterial or periarteriolar lymphoid sheath (PALS) is present around the vessels and becomes smaller toward the capillary ending. The arterioles are usually described as branching into penicillary arterioles, which run in parallel. In humans, however, this phenomenon seems to be restricted to involuted specimens in which the disappearance of tissue between arterioles has left them lying close to each other.
Branching of arterioles and capillaries often occurs at right angles, as can frequently be seen in sections. Reconstructions based on serial sections have shown that the terminal end of the capillary forms a peculiar and specifically splenic structure (25,26) (Figures 32.2,32.3). These structures are known by several names, determined partly by the species in which they have been studied, for example, sheathed capillaries, H lsekapillaren, ellipsoids, or periarteriolar macrophage sheaths. In humans, they are present in the red pulp and the perifollicular zone and are generally referred to as sheathed capillaries. The sheathed capillary is surrounded by a sheath of mononuclear phagocytes and rare reticulum cells. Because autolysis is so rapid, visualization of the sheathed capillaries in particular is dependent on adequate tissue processing. The endothelial lining of the capillary ends abruptly in a string of concentrically arranged macrophages. Blood cells coming from an arteriole have to pass through the sheathed capillary on their way to the lumen of the sinus, which they reach by slowly percolating through the cord macrophages and red pulp stroma (open circulation), and then via the slits in the basement membrane of the sinus (3,27). Although no direct anatomic connection between the arteriolar ends and the sinuses has been demonstrated, a proportion of the arteriolar branches may end in close opposition to walls of the sinuses, allowing a more rapid circulation (close circulation) of at least a portion of the blood flow. The red pulp sinuses are considered as the first part of the splenic venous tree.
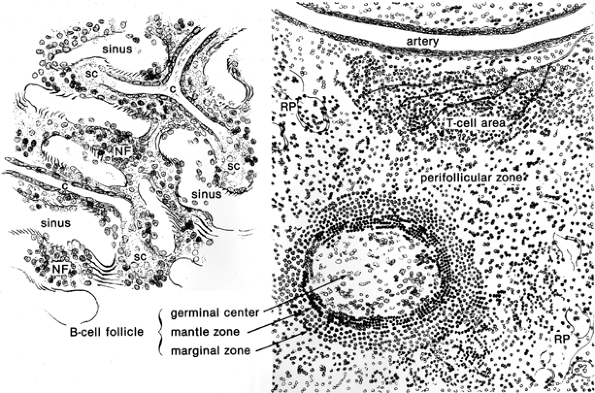 |
Figure 32.2 Schematic impression of red (left) and white (right) pulp, showing the main compartments and structures of the human spleen. The capillaries (c) end as sheathed capillaries (sc) without direct communication with the sinuses. The nonfiltering areas (NF) are bordered by sinuses and are devoid of (sheathed) capillaries. The perifollicular zone surrounds the white pulp (follicle and T-cell area) and lacks fully developed sinuses. Note the zoning in the B-cell but not in the T-cell compartment. The T-cell area contains a lymphatic plexus. (Left, original magnification 250; right, original magnification 100.) |
The localization of the sheathed capillaries at the end of the arterial tree seem perfect for their functioning as a filtering unit.
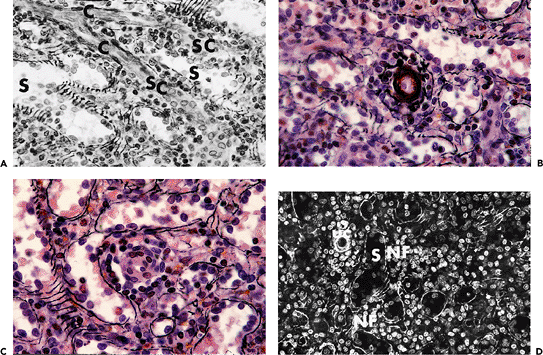 |
Figure 32.3 A. Traumatically ruptured spleen. Methylmethacrylate embedding (methenamine-silver/H&E, original magnification 400). Capillary transitioning into sheathed capillary. Note the proximity to, but lack of connection with, the sinuses. (C, capillary (unsheathed); SC, sheathed capillary; S, sinus) B. Same specimen as in A (original magnification 1000). Detail of unsheathed capillary. C. Same specimen as in A (original magnification 1000). Detail of sheathed capillary. D. Same specimen as in A (original magnification 250). Detail of the red pulp showing sinuses in cord tissue. Note the nonfiltering areas devoid of capillaries and completely surrounded by sinuses. (Uc, unsheathed capillary; S, sinus; NF, nonfiltering area) |
P.788
Within the sinusoidal meshwork, there are large sinuses that open directly into veins running along the arteries in the collagenous cuff.
Small efferent lymph vessels can be found in the T-lymphocyte compartment of the white pulp in about two-thirds of the spleens. They are not seen in the surrounding perifollicular zone. A reconstruction from serial sections showed that these lymph vessels form a network around arterioles and eventually follow the arterial tree to the hilar region (9,28).
Red Pulp
Seventy-five percent of the volume of the spleen is made up of red pulp (26). The two-dimensional picture given by conventional histology sections suggests that the red pulp is largely composed of cordal macrophages, interconnected by their cytoplasmic processes to form a reticular meshwork that provides structural support to the venous sinuses. Serial sections have shown, however, that the red pulp contains also a loose reticular framework, is rich in capillaries, and contains the terminal ends of the penicillary arterioles (Figure 32.3D). The sinuses account for about 30% of the red pulp (26). The sinus endothelial cells are surrounded by almost circular strands of discontinuous basement membrane that is predominantly composed of collagen IV and laminin, known as the ring fibers (Figure 32.4). The ring fibers are both interconnected among themselves and anchored to the dendritic processes of the cordal macrophages and splenic (fibroblastic) reticulum cells. Stromal fibers and reticulum cells running throughout the red pulp cords also contribute to provide structural support to this splenic area (the reticular meshwork of the red pulp).
P.789
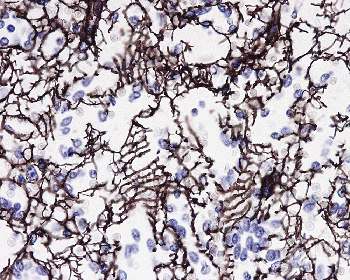 |
Figure 32.4 Normal spleen with red pulp stained with an antibody against collagen IV showing the ring fibers surrounding the sinuses (original magnification 250). |
A subpopulation of reticulum cells that express nerve growth factor receptor is found predominantly in periarteriolar location (29). These cells, most likely representing adventitial reticulum cells similar to those present in the adventitia of blood vessels, also have been observed within the stroma of bone marrow and lymph nodes (30). Myoid reticulum cells (smooth muscle actin positive, or SMA-positive) are found scattered throughout the red pulp. These cells are, however, much more concentrated within the marginal zone of the lymphoid follicles and in the PALS (31,32,33). Whether the SMA-positive red pulp cells correspond to fibroblastic reticulum cells that have undergone myofibroblastic differentiation, or are a truly separate population, is unclear at this time. The red pulp sinuses themselves form a complex network of their own with many interconnections and bulblike extensions with blind ends, the latter of which project into the cord tissue [see Figure 4 of van Krieken et al. (26)].
The sinuses are lined by elongated, flat endothelial cells with typical bean-shaped nuclei having a longitudinal cleft; these cells are also known as littoral cells. Immunoperoxidase studies have shown positivity for endothelial markers and, unique among other endothelial cells, to CD8 (Figure 32.5) and often CD68 and CD21. Parts of the endothelial lining react also with a monoclonal antibody that also immunostains high endothelial venules in the lymph node (van Krieken JHJM, unpublished observation).
The preponderant function of red pulp is blood filtration. However, in serial sections, one might notice that a fair amount of the red pulp tissue does not include capillary endings, including sheathed capillaries, and that these areas are surrounded only by sinuses. Small aggregates of lymphocytes (both B and T) and mononuclear phagocytes are present (Figure 32.3), which means that these nonfiltering areas of the red pulp should be regarded as a previously unrecognized splenic lymphoid compartment in addition to the white pulp. Morphometrically, the size of this lymphoid, nonfiltering red pulp compartment seems to be comparable to that of the white pulp (26). Newly formed white pulp follicles might originate from the small lymphoid aggregates of these nonfiltering areas.
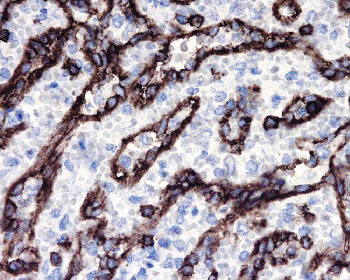 |
Figure 32.5 Normal spleen, red pulp stained with anti-CD8 showing the positive sinus endothelial cells (original magnification 400). |
Blood cells can only reach these areas by passing through large stretches of red pulp tissue or, which seems more likely, via influx from the sinus by passing through the sinus endothelium. A retrograde return of lymphocytes from the venous sinus lumen back into the splenic tissue is known for the rat spleen, where lymphocytes migrate through the walls of what is called the marginal sinus into the white pulp. This type of sinus is histologically not discernable in the white pulp of the human spleen. In humans, the role played in the rat by the marginal sinus in the exchange of lymphocytes between the sinusoidal circulation and the splenic lymphoid compartment might be played by the previously described blind-ended bulblike extensions of the red pulp sinuses, representing a splenic endothelial component with high endothelial venulelike characteristics. This hypothesis is supported by the observation that, in humans, splenic follicles are surrounded by a perifollicular zone, a distinct splenic compartment containing erythrocyte-filled vascular spaces. The perifollicular zone sinuses differ from the typical red pulp sinuses in their enhanced expression of CD34. Recent evidence has suggested that this zone may represent the entry compartment for recirculating lymphocytes into the white pulp since it is capable of supporting influx and local proliferation of lymphoreticular cells, particularly CD4-positive T lymphocytes (33).
P.790
It has been recently suggested that the entry of these cells may be dependent on the presence in the perifollicular area of specialized reticulum cells with an endothelial-like phenotype secreting lymphokines and guiding the T cells into the PALS (33).
White Pulp
The white pulp consists of B- and T-cell lymphoid compartments (11) (Figure 32.1). The B-cell compartment mainly consists of the splenic lymphoid follicles. These are composed of a germinal center (only found in secondary follicles) directly surrounded by a ring of small lymphocytes, called the mantle zone or corona, which in turn is surrounded by the marginal zone that contains medium-sized lymphocytes (Figure 32.6). The germinal centers have similar features to those found in other lymphoid organs. They are formed by a scaffold of follicular dendritic cells that express CD21, CD23, CD35, and the low-affinity nerve growth factor receptor. The B cells of the germinal centers express CD20, CD19, CD10, and CD79a but not CD5. They have a high proliferation activity with Ki-67 and do not express Bcl-2. The T lymphocytes present within germinal centers are predominantly CD4-positive; tingible-body macrophages are CD68-positive. Mantle zones consist predominantly of CD5-positive small lymphocytes that are IgM-, IgD-, Ki-B3 , and DBA.44-positive and alkaline phosphatase negative (Figure 32.7). The marginal zone lymphocytes are, in contrast, positive with alkaline phosphatase and are IgD-, Ki-B3 , and DBA.44-negative (34). The marginal zone also contains a population of macrophages that are functionally distinct from the cord histiocytes of the red pulp, which, at least in animal models, seem to be important in maintaining the anatomic structure of the marginal zone by attracting newly differentiated marginal zone B lymphocytes into it. These cells move into the marginal zone area from the germinal center, where they derive from a common follicular/marginal zone precursor B cell (35).
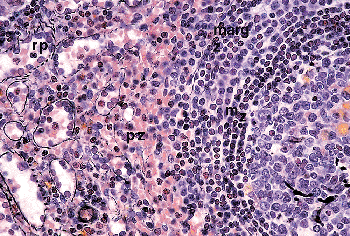 |
Figure 32.6 Same specimen as in Figure 32.3. A secondary follicle (germinal center to the right) borders the red pulp. (mz, mantle zone; margz, marginal zone; pz, perifollicular zone; rp, red pulp; original magnification 100) |
The reticulin framework of the marginal zone is characterized by the presence of numerous SMA-positive reticulum cells arranged in a concentric meshwork pattern. The marginal zone SMA-positive cells continue into the T-cell zones, where reticulum cells, exhibiting the same immunophenotype, form the reticular framework of the PALS. Cells with SMA positivity are also seen, although less frequently, in the perifollicular zone and throughout the red pulp. These cells become more prominent in the presence of red pulp congestion, such as seen in cases of fibrocongestive splenomegaly (Figure 32.8) (29,36). Contractile capability may be important for a scaffoldlike reticular structure capable of being reversibly stretched around lymphoid follicles as they expand and contract within the PALS.
In the rat spleen, the mantle and marginal zones are separated by a marginal sinus that can easily be seen by light microscopic examination. It plays an essential role in the splenic immune function as the site of entry of lymphocytes and antigens (37). This dividing sinus is not discernible in humans, at least by light microscopy. Recently, by using electron microscopy, a marginal sinuslike structure was described (38), although, surprisingly, it seems to be absent in active follicles (39). However, neither the exact location nor functional properties of this structure are known.
The light microscopic differences with rodent spleens has led to confusion in the definition of follicular structures
P.791
of the human spleen. The term marginal zone has been used with different meanings (11,40,41,42). Some investigators use the term to refer to the ring of medium-sized lymphocytes that surrounds the outer border of the mantle zone; few others have included the mantle zone, still others only the bordering area between the red and white pulp, and sometimes even the zone surrounding the T-cell areas (PALS). We prefer to reserve the term marginal zone for the unique splenic structure that encases from the outside the IgD- and IgM-positive small lymphocytes of the mantle zone (in the secondary follicle) or of the primary follicle. We refer to the bordering area between the red and white pulp as the perifollicular zone. The same definitions are used in the extensive Japanese literature on the histology of the human spleen. However, the Japanese investigators call our marginal zone the inner marginal zone and refer to the perifollicular zone as the outer marginal zone. Because of the totally different architecture and cell population of these two structures, we find it preferable to use different names.
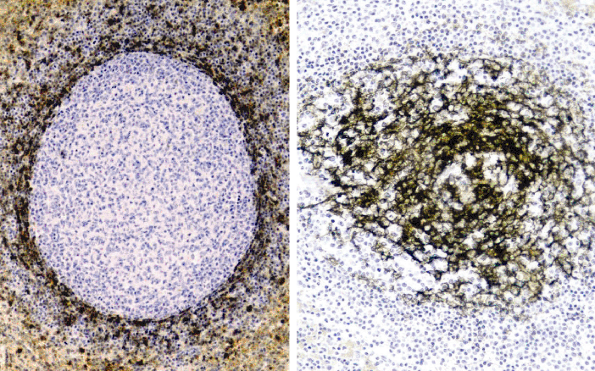 |
Figure 32.7 Normal spleen showing the positive mantle zone cells for DBA.44 (left, original magnification 40) and the dendritic cells stained for CD21 (right, original magnification 40). |
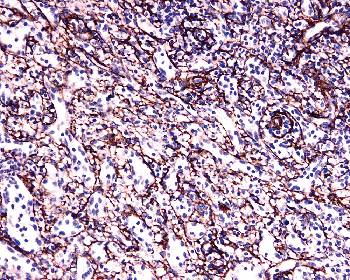 |
Figure 32.8 Spleen in fibrocongestive splenomegaly showing increased expression for smooth muscle actin (original magnification 100). |
The T-cell areas lie around arterioles but are not as regularly arranged as in the PALS seen in the rodent spleen (Figure 32.9). The arterioles are not constantly covered by these cylindrical lymphoid cuffs; they can be seen naked traversing follicles and even germinal centers.
In humans, the PALS are rather irregular aggregates of small polymorphic T lymphocytes, most of which express CD4 reactivity (T-helper/inducer lymphocytes). The T-cell areas are also surrounded by a perifollicular-like zone. The follicles sometimes border T-cell areas, with which they share a common perifollicular zone.
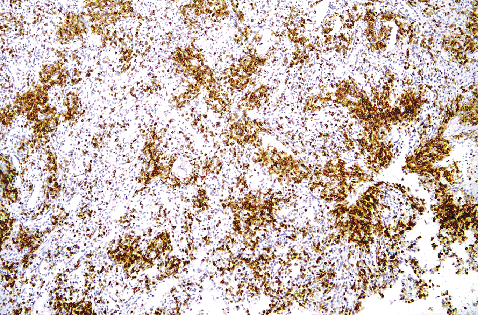 |
Figure 32.9 Normal spleen stained with anti-CD3 antibody (original magnification 40) showing the somewhat loosely organized T-cell compartment. |
P.792
Perifollicular Zone
The perifollicular zone (PFZ) is a specialized compartment of the red pulp that is associated with its own reticular stroma; PFZ is found both at the outside of the white pulp marginal zone (in the follicles) and at the periphery of the T-cell areas (PALS). In the PFZ, the reticular fibers are more widely spaced than in the rest of the red pulp (Figures 32.2 and 32.6). In silver-stained plastic sections, the PFZ can be identified by the paucity of basal membrane strands and by the presence of a vascular pattern that is different from the one seen in the rest of the red pulp. At the outer border of this area, red pulp sinuses are more widely spaced than elsewhere, and a rich network of capillaries, including sheathed capillaries, is present. The PFZ contains a considerable numbers of erythrocytes and leukocytes.
The PFZ zone at the outside of the white pulp stands out in silver-stained sections but may be poorly visible in routine hematoxylin- and eosin (H&E)-stained sections. However, since the PFZ contains a large number of erythrocytes, it can be recognized by its deeply congested appearance around the densely packed lymphocytes of the T- and B-cell areas (Figure 32.1A). The erythrocytes are seen more regularly in PFZ than in the red pulp sinuses; the red pulp sinuses outside the PFZ often appear to be less filled with these cells than the cord tissue.
The PFZ, which makes up about 8% of the spleen, contains a mixture of blood cells comparable with that of the peripheral blood. It has been suggested that this area is responsible for the passage of the about 10% of the splenic blood, which is known to have a retarded flow (11).
Flow Cytometry
The spleen has been only rarely investigated by flow cytometry. Recently, we have looked at the immunophenotypic characteristics of the human spleen lymphocytes and have established reference values for those cells in normal and reactive spleens (43). Altough the results obtained showed B- and T-antigen patterns of expression comparable to those seen in other lymphoid organs, a few differences were noted (Table 32.2). In contrast to the thymus and bone marrow, the spleen contains only very rare Tdt-positive lymphoid precursors. Within the Bcell subsets, the spleen shows a frequency of CD19-positive/CD20-negative B cells that is higher than in the peripheral blood or lymph node. This corresponds to the presence, in the spleen, of a sizable proportion of early plasma cells (CD138-negative) as well as more mature plasma cells. Other findings included a significant population of CD20/CD5 positive B cells, accounting for approximately 10% of the B lymphocytes; the presence of CD4/CD8 ratio of 1.2:1 (lower than in the blood but similar to the one seen in lymph node) and a mean number of / -positive T cells (of all CD3-positive cells) of 6% in normal and 10% in reactive spleens. While in the peripheral blood, NK/T cells account for less than 6% of the CD3-positive circulating lymphocytes, our study showed a relatively high frequency of these cells in the spleen. Similar results have been observed in the liver by others and support the concept that spleen and liver lymphocytes have specific, although yet unknown, immunoregulatory functions (43).
Ultrastructure
Electron microscopy (especially scanning electron microscopy), including the use of microcasts from the vasculature, has elucidated largely the functional microanatomy of the spleen. These studies have shown the routes that blood cells take through the spleen and have also clearly illustrated the pitting function of the spleen (removal of inclusions in erythrocytes) exerted by the sinusoids. In spleen examined in a diagnostic-oriented setting, however, there is hardly if ever the necessity of using ultrastructural studies.
Function
The human spleen has several important functions. However, splenectomy in general does not lead to impaired health, except for an increased risk of overwhelming postsplenectomy infections caused by encapsulated bacteria (e.g., pneumococci). The reason for this is that many functions of the spleen, at least in adults, can be taken over by other organs.
In humans, the spleen is involved in the primary immune response to bloodborne antigens and polysaccharide antigens; it also acts as a regulator of immune reactions elsewhere in the body. It contains a specific environment that facilitates the binding of antibodies and antigens; cells or microorganisms covered by antibodies are trapped and destroyed in the spleen, as are erythrocytes that have decreased flexibility and lowered osmotic resistance (1,44,45,46,47,48,49,50).
P.793
Each of these functions takes place in a specific splenic compartment, which is capable of undergoing rapid changes in its size and composition, even under physiologic conditions. Therefore, the main splenic functions to be considered include blood filtering, immunologic function, hematopoiesis, and reservoir.
Table 32.2 Expression of Surface and Intracellular Markers by Human Spleen Lymphocytes | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
Filter Function
The location and specialized anatomy of the spleen is especially suitable for its function as a filter of the blood. Normal blood cells are capable of traversing the barrier of macrophages of the sheathed capillary, the red pulp cord macrophages, and the sinus endothelium (collectively, the filtering unit of the spleen) at a speed comparable to that of the blood in the capillary bed of other organs. However, in cases in which the flexibility of the red blood cell is diminished (e.g., by aging, intoxication, or congenital defects), the macrophages of the splenic filtering unit can eliminate the abnormal cell by ingesting it. The filtering function includes a process known as pitting, a term which is used to describe the removal of inclusions, such as nuclear remnants known as Howell-Jolly (H-J) bodies, from erythrocytes without destroying the cell. The presence of H-J bodies in circulating erythrocytes in the peripheral blood indicates the presence of splenic hypofunction (e.g., splenectomized patients).
P.794
In addition to red blood cells, the macrophages of the spleen can readily take up bacteria, antigens, and immune complexes. The spleen is capable of filtering out reticulocytes, platelets, hematopoietic stem cells, lymphocytes, and dendritic cells from the blood and provides the proper microambient conditions for their further differentiation. Also, it sequesters monocytes from the blood and facilitates their transformation into splenic macrophages (8).
Immunologic Function
The spleen plays a more important role in the development of the immune system, but even in adults the spleen is still involved in B- and T-lymphocyte production and differentiation. The spleen receives B and T cells from the recirculating lymphocyte pool and sorts them into dedicated compartments such as the follicles and the PALS, where they can interact with antigens and antigen-presenting cells and become capable of mounting effective immune responses.
The marginal zone is a component of the B-cell follicle and is a remarkably larger compartment in the spleen than elsewhere (e.g., tonsils). Although the exact physiologic function(s) of the marginal zone is still unclear (42), its main immunologic role relates to the thymus-independent rapid response to bloodborne microorganisms; since these are rapidly trapped in the spleen and brought directly into contact with numerous immunocompetent cells, the spleen is well situated for this task.
Hematopoiesis
In rodents, the spleen has a large hematopoietic function, but this is not the case in humans. As described above, the hematopoietic function is only present in the fetal spleen; in the adult spleen, hematopoiesis does not occur (15). Hematopoietic cells encountered in the adult spleen originate from circulating, marrow-derived, progenitors/early precursors that become entrapped in the spleen and are capable of undergoing further differentiation. When this physiologic phenomenon reaches pathologic relevance by causing splenomegaly, it is termed splenic myeloid metaplasia or extramedullary hematopoiesis. Although splenic myeloid metaplasia can be seen in many different conditions, the most striking examples of this condition can be observed in patients with chronic idiopathic myelofibrosis (15).
Reservoir Function
The human spleen contains about 300 mL of blood. This is a relatively small amount, in contrast to that seen in a dog or cat. In these animals, the spleen functions as an important blood reservoir; and in situations where more blood is needed, its rapid contraction can increase substantially the amount of circulating blood cells. It is highly doubtful that this function occurs at all in humans, whose splenic capsule lacks a significant component of smooth muscle fibers. The spleen, however, does function as a reservoir for factor VIII of the clotting system, platelets, granulocytes, and iron.
Aging Differences
In infancy and childhood, the immune system is not yet fully developed, and this is also reflected in the histology of the spleen. The marginal zone is observed as a separate compartment only after four months of age; moreover, the marginal zone B cells in the spleen of infants have a different phenotype (lack CD21; IgD- and IgM-positive) compared with adult marginal zone B cells (51). An important age difference, in our experience, is the regular occurrence of germinal centers in the white pulp of normal spleens in patients younger than 20 years; older patients have been shown to have only rare secondary follicles (9). The often mentioned age-dependent atrophic change has only been documented in patients in their eighth decade of life (11).
Hyalinization of vessels in the spleen is seen frequently, even in very young children and, therefore, does not represent a pathologic finding (52).
Differential Diagnosis
In the spleen, compartmentalized lymphoid tissue (white pulp) is interwoven by the filtering red pulp. Each splenic compartment reacts to external stimuli with physiologic changes in its composition and histology. As in the lymph node, the line between pathologic and impressive but essential physiologic reactions is vague. The amount of white pulp, for instance, varied from 5 to 22% of the total splenic tissue in a normal control group (11).
As previously mentioned, normal blood cells can pass undamaged through the barrier of macrophages of the sheathed capillary and the red pulp cord tissue as well as the sinus endothelium (i.e., the filtering unit of the spleen) at a speed comparable with that of the blood flow in the capillary bed of other organs. However, when the flexibility of the blood cells is diminished (e.g., by aging, intoxication, or congenital defects), the red pulp macrophages can ingest the abnormal cells. In this process, the sheathed capillaries seem to lose their macrophages, which spread out into the surrounding red pulp or enter the sinuses to be transported to the liver. In cases characterized by chronic stimulation of the filtering function, it can be demonstrated that the amount and length of the capillaries increase in parallel with the hypertrophy of the red pulp, whereas the sheathed capillaries are less readily seen in the sections (26). In idiopathic thrombocytopenic purpura (ITP), remnants of
P.795
phagocytosed thrombocytes can be seen as periodic acid-Schiff (PAS)-positive fragments in cord macrophages. If blood cells are covered by immunoglobulins or immunocomplexes, parts of the cell membrane can be removed by the sinus endothelium by pitting and culling, giving rise to a spherocyte. This happens in a fashion similar to the removal of nuclear remnants, as previously described (15).
In septicemia, the filtering compartment may show morphologic findings (activation and hyperplasia of macrophages) indistinguishable from those seen in cases of acute or chronic hemolysis; these changes are most likely induced by the presence of circulating immunocomplexes, fragmented cells, or antibody-coated cells. In these conditions, postmortem autolysis of the activated macrophages can lead to early disintegration of the red pulp cells and stroma. The septic spleen at autopsy thus probably represents an artifact that can be the result of, but is not specific for, sepsis; it, especially, should not be diagnosed as splenitis. True splenitis, in which the spleen contains an inflammatory response to a local noxious agent such as in typhoid fever or tropical diseases, is rare in the Western hemisphere. Lymphoplasmacytoid cells and plasma cells normally rim arteries and arterioles and extend along red pulp capillaries. This perivascular cellular rim also may contain some macrophages or small epithelioid granulomas, the significance of which is unclear. The perivascular presence of plasma is a normal finding and by no means justifies a diagnosis of splenitis, nor does the diffuse influx of granulocytes throughout the red pulp in specimens resected during prolonged surgery.
The effects of chronic venous congestion are not clear. In our preliminary studies in patients dying with chronic cardiac disease, the so-called effect of chronic cardiac congestion on the lymphoid and filtering compartments appears more likely to be the effect of concomitant infections or is therapy mediated. In chronic venous congestion due to portal hypertension, the sinuses are normal in size but contain fewer buds and appear rigid. The amount of cord tissue and the number of capillaries are both decreased; in the cord tissue, an increase of reticular fibers (fibrocongestive splenomegaly) and increased expression of smooth muscle actin in reticulum cells is seen (26,36). Infarcts in the splenic tissue are microscopically more irregularly defined and poorly demarcated than could be expected macroscopically due to the intricate distribution of the splenic vessels. In three-dimensional reconstructions, capillaries from different arterioles are seen to cross each other with overlapping territories.
Primary tumors of the spleen are rare. Metastatic carcinoma seems especially to occur in neuroendocrine tumors, including small cell carcinoma of the lung, with a conspicuous tendency for intrasinusoidal spread. Malignant lymphomas exhibit a homing pattern to specific splenic compartments dependent on the type of lymphoma, similar to that observed in other lymphoid organs (21). In nonlymphomatous hematopoietic malignancies involving the spleen, their distribution pattern is similar to that observed in the bone marrow: extramedullary erythropoiesis and megakaryopoiesis are found primarily along and within the sinuses of the red pulp, whereas myelomonopoiesis is found in proximity of the capillaries within the cord tissue. Blastic infiltration seen in cases of acute leukemia can be found anywhere in the spleen.
Specimen Handling
The spleen is quite vulnerable and, due to the large numbers of macrophages and granulocytes, may undergo rapid autolysis. Proper and rapid fixation is therefore important, and this goal is not reached when the entire organ is put into formalin. For proper handling, the specimen has to be received fresh, and handling has to be rapid. The organ is weighed and the surface examined. After that, the organ is cut into small slices of 0.5 cm. Then the cut surface is inspected carefully for nodules larger than normal white pulp. Ideally pieces should be frozen for flow cytometry, cryostat section immunohistochemistry, or other techniques. When no abnormalities are seen, at least three or four blocks are taken out randomly and processed for microscopic examination.
Histologic Technique
Routine paraffin embedding leads to shrinkage and loss of cellular detail. Because routine H&E staining often does not yield sufficient information, the use of appropriate histology techniques, such as plastic embedding (53), methenamine-silver/H&E stain (54), or at least, PAS and Gomori's reticulin stains, is necessary for an adequate morphologic analysis of the splenic microarchitecture (Figure 32.10).
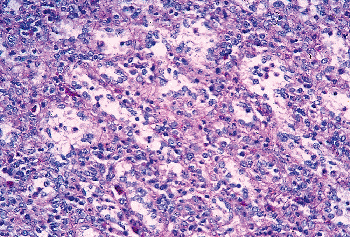 |
Figure 32.10 Same specimen as in Figure 32.1. Detail of red pulp showing with some difficulty the structure of the sinuses (PAS stain, paraffin-embedded, original magnification 200). |
P.796
Special Procedures
The spleen is only rarely removed for diagnostic purposes. Staging laparotomy is no longer part of the required diagnostic workup of a patient with Hodgkin's disease. Therefore, splenic pathology may be seen as an unsuspected incidental finding in a patient splenectomized for other reasons (e.g., chronic idiopathic thrombocytopenia, trauma). Not uncommonly, splenic lymphoma may be discovered as an incidental finding. A high degree of suspiciousness of the grossing pathologist is necessary in these case since, for optimal lymphoma diagnosis, it is often necessary to apply techniques that require fresh/frozen tissue, such as flow cytometry, frozen tissue immunohistochemistry, molecular analysis, and/or cytogenetics (Figure 32.8).
In view of the frequent lack of fresh specimens, clonality assessment in a spleen is usually done by immunohistochemistry applied to paraffin-embedded tissue, looking for a restricted pattern of immunoglobulin light chain expression in B cells. This is one of the most important special techniques used in routine diagnostic laboratories. In addition, clonality can also be established by using a polymerase chain reaction based technique for detecting immunoglobulin gene rearrangement, which can also be succesfully applied to paraffin-embedded tissue (55).
Immunohistochemistry can also be necessary to characterize other hematopoietic and nonhematopoietic tumors, the latter primary or metastatic, which can occur (although infrequently) in the spleen. In storage disorders such as Gaucher's disease, electron microscopy can be of additional value, although biochemical analysis is considered by most experts as the most practical and specific approach.
Conclusion
The human spleen has always been a somewhat enigmatic organ. Studies of its histology must be based on carefully selected, surgically excised normal spleens. The organ should be processed immediately and appropriately for optimal results.
Previous studies by one of us of a large series of spleens with adequate histologic techniques and with reconstruction based on serial sections has shown that the spleen is a highly compartmentalized organ (Table 32.1). Each compartment has its own structure and cell populations and often a separate function. The old division into red and white pulp is probably oversimplified and should be expanded.
Human and animal spleens are different in many important structural aspects; data extrapolation from animal studies to humans is therefore problematic and often unwarranted.
References
1. Crosby WH. The spleen. In: Wintrobe MM, ed. Blood, Pure and Eloquent. New York: McGraw-Hill; 1980:96 138.
2. Chen L, Weiss L. Electron microscopy of the red pulp of human spleen. Am J Anat 1972;134:425 458.
3. Weiss L. The spleen. In: Greep RO, Weiss L, eds. Histology. 3rd ed. New York: McGraw-Hill; 1973:545 573.
4. Nieuwenhuis P, Keuning FJ. Germinal centres and the origin of the B-cell system. II. Germinal centres in the rabbit spleen and popliteal lymph nodes. Immunology 1974;26:509 519.
5. Nieuwenhuis P, Ford WL. Comparative migration of B- and T-lymphocytes in the rat spleen and lymph nodes. Cell Immunol 1976;23:254 267.
6. Ford WL. Lymphocyte migration and immune responses. Prog Allergy 1975;19:1 59.
7. Veerman AJ, van Ewijk W. White pulp compartments in the spleen of rats and mice. A light and electron microscopic study of lymphoid and non-lymphoid celltypes in T- and B-areas. Cell Tissue Res 1975;156:417 441.
8. Weiss L. The spleen. In: Weiss L, ed. Cell and Tissue Biology: A Textbook of Histology. 6th ed. Baltimore: Urban & Schwarzenberg; 1988:517 538.
9. van Krieken JH, te Velde J, Kleiverda K, Leenheers-Binnendijk L, van de Velde CJ. The human spleen: a histological study in splenectomy specimens embedded in methylmethacrylate. Histopathology 1985;9:571 585.
10. Scothorne RJ. The spleen: structure and function. Histopathology 1985;9:663 669.
11. van Krieken JH, te Velde J, Hermans J, Cornelisse CJ, Welvaart K, Ferrari M. The amount of white pulp in the spleen: a morphometrical study done in methacrylate-embedded splenectomy specimens. Histopathology 1983;7:767 782.
12. van Krieken JHJM. The architecture of the human spleen. Academic thesis. Pijnacker, The Netherlands: Dutch Efficiency Bureau; 1985.
13. van Krieken JH, te Velde J. Immunohistology of the human spleen: an inventory of the localization of lymphocyte subpopulations. Histopathology 1986;10:285 294.
14. Wolf BC, Luevano E, Neiman RS. Evidence to suggest that the human fetal spleen is not a hematopoietic organ. Am J Clin Pathol 1983;80:140 144.
15. Neiman RS, Orazi A. Disorders of the Spleen. 2nd ed. Philadelphia: WB Saunders; 1999.
16. Timens W, Rozeboom T, Poppema S. Fetal and neonatal development of human spleen: an immunohistological study. Immunology 1987;60:603 609.
17. Jones JF. Development of the spleen. Lymphology 1983;16:83 89.
18. Namikawa R, Mizuno T, Matsuoka H, et al. Ontogenic development of T and B cells and non-lymphoid cells in the white pulp of human spleen. Immunology 1986;57:61 69.
19. Moller JH, Nakib A, Anderson RC, Edwards JE. Congenital cardiac disease associated with polysplenia. A developmental complex of bilateral left-sidedness. Circulation 1967;36:789 799.
20. Myers J, Segal RJ. Weight of the spleen. I. Range of normal in a nonhospital population. Arch Pathol 1974;98:33 35.
21. van Krieken JH, Feller AC, te Velde J. The distribution of non-Hodgkin's lymphoma in the lymphoid compartments of the human spleen. Am J Surg Pathol 1989;13:757 765.
22. Seufert RM. Chirurgie der Milz. Stuttgart, Germany: Enke-Verlag; 1983.
23. Tischendorf F. Blutgef ss und Lympfgef ssapparat Innersekretorische Drusen. Die Milz. Berlin: Springer-Verlag: 1969.
24. Snook T. A comparative study of the vascular arrangements in mammalian spleens. Am J Anat 1950;87:31 77.
25. Buyssens N, Paulus G, Bourgeois N. Ellipsoids in the human spleen. Virchows Arch A Pathol Anat Histopathol 1984;403:27 40.
26. van Krieken JH, te Velde J, Hermans J, Welvaart K. The splenic red pulp: a histomorphometrical study in splenectomy specimens embedded in methylmethacrylate. Histopathology 1985;9:401 416.
P.797
27. Heusermann U, Stutte HJ. Comparative histochemical and electron microscopic studies of the sinus and venous walls of the human spleen with special reference to the sinus venous connections. Cell Tissue Res 1975;163:519 533.
28. Fukuda T. Deep lymphatics of the spleen. Tohoku J Exp Med 1963;79:281 292.
29. Orazi A, O'Malley DP, Thomas JL, Czader M. Stromal changes in reactive and malignant disorders of the spleen. Mod Pathol 2004;17:264A.
30. Cattoretti G, Schiro R, Orazi A, Soligo D, Colombo MP. Bone marrow stroma in humans: anti-nerve growth factor receptor antibodies selectively stain reticular cells in vivo and in vitro. Blood 1993;81:1726 38.
31. Pinkus GS, Warhol MJ, O'Connor EM, Etheridge CL, Fujiwara K. Immunohistochemical localization of smooth muscle myosin in human spleen, lymph node, and other lymphoid tissues. Unique staining patterns in splenic white pulp and sinuses, lymphoid follicles, and certain vasculature, with ultrastructural correlations. Am J Pathol 1986;123:440 453.
32. Toccanier-Pelte MF, Skalli O, Kapanci Y, Gabbiani G. Characterization of stromal cells with myoid features in lymph nodes and spleen in normal and pathologic conditions. Am J Pathol 1987;129:109 118.
33. Steiniger B, Barth P, Hellinger A. The perifollicular and marginal zones of the human splenic white pulp: do fibroblasts guide lymphocyte immigration? Am J Pathol 2001;159:501 512.
34. van Krieken JH, von Schilling C, Kluin PM, Lennert K. Splenic marginal zone lymphocytes and related cells in the lymph node: a morphologic and immunohistochemical study. Hum Pathol 1989;20:320 325.
35. Pillai S, Cariappa A, Moran ST. Marginal Zone B lymphocytes. Annu Rev Immunol 2005;23:161 196.
36. Kraus MD. Splenic histology and histopathology: an update. Semin Diagn Pathol 2003;20:84 93.
37. Sasou S, Satodate R, Katsura S. The marginal sinus in the perifollicular region of the rat spleen. Cell Tissue Res 1976;172:195 203.
38. Schmidt EE, MacDonald IC, Groom AC. Microcirculatory pathways in normal human spleen, demonstrated by scanning electron microscopy of corrosion casts. Am J Anat 1988;181:253 266.
39. Schmidt EE, MacDonald IC, Groom AC. Changes in splenic microcirculatory pathways in chronic idiopathic thrombocytopenic purpura. Blood 1991;78:1485 1489.
40. Takasaki S. Light microscopic, scanning and transmission electron microscopic, and enzyme histochemical observations on the boundary zone between the red pulp and its surroundings in human spleens. Tokyo Yikekai Med J 1979;94:553 568.
41. Yamamoto K, Arimasa N, Yamamoto T, Tokuyama K, Kobayashi T, Itoshima T. Scanning electron microscopy of the perimarginal cavernous sinus plexus of the human spleen. Scan Electron Microsc 1979;3:763 768.
42. Kraal G. Cells in the marginal zone of the spleen. Int Rec Cytol 1992;132:31 74.
43. Colovai AI, Giatzikis C, Ho EK, et al. Flow cytometric analysis of normal and reactive spleen. Mod Pathol 2004;17:918 927.
44. Koyama S, Aoki S, Deguchi D. Electron microscopic observations of the splenic red pulp with special reference to the pitting function. Mie Med J 1964;14:143 188.
45. Sampson D, Grotelueschen C, Kauffman HM Jr. The human splenic suppressor cell. Transplantation 1975;20:362 367.
46. Videbaok A, Christensen BE, Jonsson V. The Spleen in Health and Disease. Chicago: Year Book Medical; 1983.
47. Wyler DJ. The spleen in malaria. In: Evered D, Whelan J, eds. Malaria and the Red Cell. London: Pitman; 1983.
48. Van Krieken JH, Breedveld FC, te Velde J. The spleen in Felty's syndrome: a histological, morphometrical, and immunohistochemical study. Eur J Haematol 1988;40:58 64.
49. Claassen E. Histological organization of the spleen: implications for immune functions in different species (38th forum in immunology). Res Immunol 1991;142:315 372.
50. van Rooijen N, Claassen E, Kraal G, Dijkstra C. Cytological basis of immune functions of the spleen. Immunocytochemical characterization of lymphoid and non-lymphoid cells involved in the in situ immune response. Prog Histochem Cytochem 1989; 19:1 71.
51. Timens W, Boes A, Rozeboom-Uiterwijk T, Poppema S. Immaturity of the human splenic marginal zone in infancy. Possible contribution to the deficient infant immune response. J Immunol 1989;143:3200 3206.
52. Lindley RP. Splenic arteriolar hyalin in children. J Pathol 1986;148:321 325.
53. te Velde J, Burkhardt R, Kleiverda K, Leenheers-Binnendijk L, Sommerfeld W. Methyl-methacrylate as an embedding medium in histopathology. Histopathology 1977;1:319 330.
54. Jones DB. Nephrotic glomerulonephritis. Am J Pathol 1957;33:313 329.
55. van Dongen JJ, Langerak AW, Bruggemann M, et al. Design and standardization of PCR primers and protocols for detection of clonal immunoglobulin and T-cell receptor gene recombinations in suspect lymphoproliferations: report of the BIOMED-2 Concerted Action BMH4-CT98-3936. Leukemia 2003;17:2257 2317.