16 - Larynx and Pharynx
Editors: Mills, Stacey E.
Title: Histology for Pathologists, 3rd Edition
Copyright 2007 Lippincott Williams & Wilkins
> Table of Contents > VI - Thorax and Serous Membranes > 21 - Serous Membranes
21
Serous Membranes
Darryl Carter
Lawrence True
Christopher N. Otis
Anatomy
The mesothelium lines the pleural, pericardial, and peritoneal cavities. Mesothelial cells on the serous surfaces appear as a simple or cuboidal epithelium, although they are of mesodermal origin. They are supported by a fibrous submesothelial layer, which becomes continuous with the outer layer of invested viscera. The serous membranes show functional differentiation according to their derivation from visceral or parietal mesoderm.
Because of space limitations, description of the gross anatomy of the mesothelium must be somewhat truncated, but some areas have functional differentiation that is reflected by their histologic features. The pleura is a continuous membrane that covers the chest wall and the lungs. The visceral pleura coats the entire pulmonary surface, including the major and minor fissures that divide the lung into lobes, whereas the parietal pleura extends over the ribs, sternum, and supporting structures and is reflected over the mediastinal structures on either side. Posteriorly in the mediastinum, the two layers of parietal pleura are separated by a thin band of fibrovascular connective tissue. Superiorly, the cervical pleura is reflected into the retroclavicular area over the apex of the lung and is coated by a thickened layer of fibrous tissue and skeletal muscle; inferiorly, the diaphragmatic pleura represents its caudal extent. Anteriorly, the pleura is reflected over part of the pericardium. The posterior visceral pleura becomes continuous with the diaphragmatic pleura over the pulmonary ligament. The heart and great vessels lie in the pericardium, which is lined by a continuous layer of mesothelium. The visceral (epicardial) side is connected to the myocardium, and the parietal (pericardial) layer rests on a dense fibrous tissue layer containing branches of the internal mammary and musculophrenic vessels, descending aorta, and branches of the vagus, phrenic, and sympathetic nerves. The thoracic surface of the pericardium is coated with parietal pleura.
The peritoneum is a nearly continuous membrane lining the potential space between the intra-abdominal viscera and the abdominal wall. In the female, it is normally interrupted by the lumina of the fallopian tubes. Anatomically, it is more complex than either the pleura or pericardium. The parietal layer covers the abdominal wall, diaphragm, anterior surfaces of the retroperitoneal viscera, and the pelvis. The visceral peritoneum invests the intestines and other intra-abdominal viscera. The elongated structures in which the parietal and visceral layers come together are the mesentery, which contains blood vessels, lymphatics, lymph nodes, and nerves.
The greater omentum is a double sheet with four layers of mesothelium between which there are numerous blood vessels and adipose tissue, which may be abundant; lymphatics and lymph nodes are less prominent than in the
P.548
mesentery. The peritoneal cavity is grossly divided into the greater sac over the intestines, the retrogastric lesser sac, right and left retrocolic areas, and the pelvis. Several outpouchings of peritoneum are often seen in pathology laboratories. Inguinal hernia sacs are pouches of parietal peritoneum, often invested with fibrous tissue and occasionally with skeletal muscle, which have been pushed through the abdominal musculature into the inguinal canal. Umbilical or ventral hernias are also outpouchings of peritoneum, but the specimens received by pathologists after surgery for their repair are usually preperitoneal fibroadipose tissue pushed ahead of the parietal peritoneum rather than mesothelium itself.
The scrotum acquires a lining of parietal mesothelium, the processus vaginalis, into which the testes descend during the seventh month of gestation. A mesothelial layer forms the surface of the tunica vaginalis. Distention of this mesothelial sac on the tunica vaginalis results in a hydrocele communicating with the peritoneal cavity when congenital but noncommunicating in acquired hydroceles. The sac of an inguinal hernia communicates with the peritoneal cavity and not with the mesothelium-lined space of the scrotum. Both hernia and hydrocele sacs are capable of a wide range of reactive changes.
Functional Anatomy
The functional anatomy of the pleura has been described by Sahn (1) and Pistolesi et al. (2). The pleura is a continuous membrane surrounding a space that normally contains approximately 10 mL of clear colorless fluid. The surface is lined by a single layer of mesothelial cells anchored to a basement membrane that lies on layers of collagen and elastic tissue containing vascular and lymphatic vessels. The lining mesothelial cells are 16 to 40 m in diameter, have rounded nuclei, usually containing a nucleolus, and a relatively large amount of cytoplasm. Although the visceral and parietal pleurae are opposing parts of the same continuous membrane, there are major functional differences between them.
The human visceral pleura is thick relative to that seen in some other mammals (3) and is similar to that of horses, cattle, sheep, and pigs (4). It has an arterial blood supply from the bronchial arteries, with a venous return that passes first into the pulmonary veins and then into the left atrium except for certain hilar regions that are drained by bronchial veins into the right atrium. The lymphatics that pass through the visceral pleura are the superficial layer of pulmonary lymphatics with extensive connections to the peribronchial, perivascular, and interlobular lymphatic spaces and lymphoid tissue (5). Blood and lymphatic vessels are invested by collagen and elastic fibers, which are divided into two layers: an external elastic lamina supporting the mesothelial cells and an internal layer investing the vessels and becoming continuous with the pulmonary interstitium (Figures 21.1,21.2). Histologic identification of the layer of elastin has been considered clinically important in determining pleural invasion, which is significant for staging of primary lung cancer (6), but elastin may also be interrupted in nonneoplastic conditions that scar the pleura. In sheep, and probably in humans, the thickness of the external layer increases in both craniocaudal and ventrodorsal directions (7). The visceral pleura is innervated by branches of the vagus nerves and sympathetic nerve trunks.
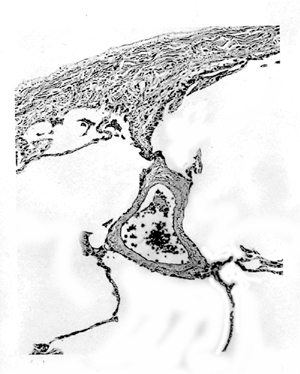 |
Figure 21.1 Visceral pleura. The mesothelial cells on the surface are thin, in profile, and not apparent. The dense submesothelial layer from the posterior surface of the left lower lobe is composed of collagen and elastin. It extends into adjacent pulmonary interstitium and around pulmonary vessels. |
The parietal pleura is anatomically, histologically, and functionally different. Although the single layer of mesothelial cells that lie on the surface of the parietal pleura are cytologically similar to those that form the continuous membrane over the visceral pleura, they are interrupted by stomata which range in size from 2 to 12 m in diameter (8,9,10,11,12). The stomata communicate directly with lymphatic lacunae that are surrounded by bundles of collagen and drain directly into intercostal lymphatics and then into the mediastinum, where they are particularly dense along the retrocardiac surface (13,14,15,16). Fluid and particulate matter extravasated from the lung are collected in these lymphatics and passed into the mediastinum, where the mesothelium covers collections of macrophages called Kampmeier foci (17,18). The arterial and venous blood supply to the parietal pleura is from the intercostal
P.549
vessels. The thickness of the fibroelastic layer investing the parietal pleural lymphatics is relatively constant and considerably less than that of most of the visceral pleura, suggesting that it serves as a membrane across which fluid may diffuse. The parietal pleura is innervated by branches of the intercostal nerves, which are activated in pleurisy. The structures of the peritoneum are similar to those of the pleura (19,20).
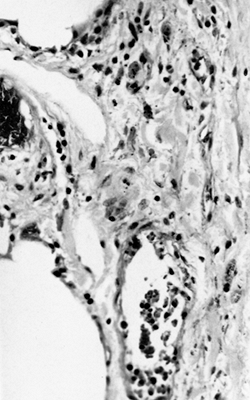 |
Figure 21.2 Visceral pleura. Capillaries are prominent. The lymphatics are dilated but deeply placed and entirely invested by the submesothelial layer. |
Functions of Serous Membranes
The serous membranes serve as selective barrier for fluid and cells. A small volume of fluid is required for capillary action to facilitate adherence of visceral and parietal pleurae as the lungs and chest wall expand and contract. Elements of the serous membranes regulate fluid interchange to keep this fluid at a mimimal level to prevent compromise of the lung volumes. Control appears to be at the capillary level because fluid is freely diffusible through mesothelium and submesothelial stroma. Another level of control results in the relatively low protein content (1.0 to 1.5 g/dL) of pleural fluid. The point of protein regulation is unknown, although speculation is that it is at the level of mesothelial microvilli (21). In the thoracic cavity, the direction of flow appears to be via diffusion from capillaries of both visceral and parietal pleurae, with resorption primarily through parietal pleural capillaries. Turnover is estimated at 0.7 mL/hr (21) (Figure 21.3). Small molecules (less than 4 nm in diameter) diffuse through the intercellular spaces and junctions between mesothelial cells. Loss of control results in serous effusions such as those seen in congestive heart failure.
Larger molecules, up to 50 nm in diameter, are transferred across the mesothelium by pinocytotic uptake and transcellular transport. Larger structures, such as cells in bloody effusions, are transported via the stomata and crevices. Loss of control of these mechanisms results in accumulations of exudative pleural fluid. Mesothelial cells express the secretory component of IgA, which is otherwise limited to surfaces with direct environmental contact (22). The glycoprotein-rich pleural fluid acts as a lubricant to minimize friction between visceral and parietal pleurae. The site of synthesis and mechanisms of control of the carbohydrate-rich fractions of the pleural fluid are unknown. The submesothelial connective tissue distributes mechanical
P.550
forces from the pleura uniformly throughout the lungs. Such a redistribution of forces is not required of the abdominal serosa. Both mesothelial cells and fibroblasts contribute to collagen synthesis.
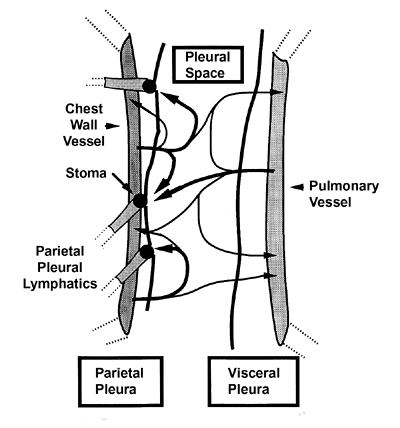 |
Figure 21.3 Model of the dynamics of pleural fluid formation. A transudate from capillaries in visceral and parietal pleura is partly reabsorbed by those capillaries and the rest diffuses into the pleural space, where it is resorbed via stomata into parietal pleural lymphatics. |
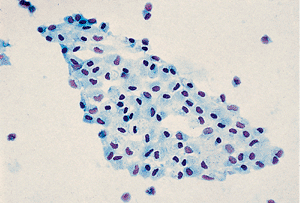 |
Figure 21.4 In this peritoneal wash specimen, a sheet of normal mesothelial cells has been detached. |
Mesothelial Cells
Morphology
Normal mesothelial cells are rarely seen in histologic sections but may be evident in cytologic preparations of peritoneal washes taken during a laparotomy (Figure 21.4). When thus visualized, they have abundant clear cytoplasm with crisply defined cell borders, small and centrally placed nuclei with a homogeneous chromatin pattern, and usually without a nucleolus (Figure 21.5).
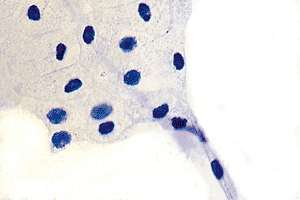 |
Figure 21.5 At higher magnification, a sheet of relatively normal mesothelial cells with abundant, clear cytoplasm and crisply defined cell borders. The centrally placed nuclei are small and have a homogenous chromatin. |
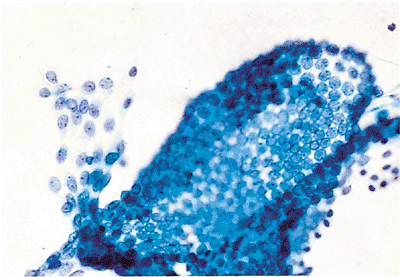 |
Figure 21.6 This detached fragment of reactive mesothelium shows an intact mesothelial layer with cells in two phases of the reactive process shown in Figures 21.7 and 21.8. |
In a variety of reactive processes, the mesothelial cells undergo markedly proliferative and hyperplastic changes. A relatively abundant cytoplasm is maintained, but the cell borders are less sharply defined. The nuclei are larger, both absolutely and relatively, the chromatin pattern is more hyperchromatic, and nucleoli are often present and prominent (Figures 21.6,21.7,21.8,21.9).
As the hyperplastic changes in the reactive mesothelial cells progress, cell groups become smaller, and individual cells predominate. When clustered, reactive mesothelial cells present an irregular outside border. The nucleus, and especially the nucleolus, may enlarge dramatically, but the nuclei are similar in size, shape, and pattern from cell to cell. Mitotic figures may be seen. The cytoplasm may become multivacuolated as the cells degenerate and imbibe fluid (Figures 21.10,21.11,21.12,21.13,21.14,21.15,21.16,21.17,21.18).
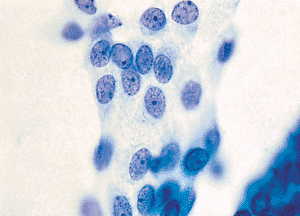 |
Figure 21.7 These reactive mesothelial cells from the left side of Figure 21.6 have abundant cytoplasm, and the nuclei are larger with a more vesicular chromatin pattern. Nucleoli are present but not prominent. |
P.551
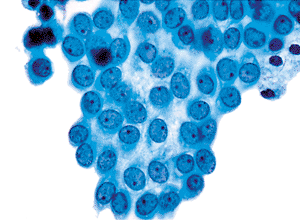 |
Figure 21.8 The more reactive mesothelial cells from the right side of Figure 21.6 have larger nuclei, a more vesicular chromatin pattern, and more prominently featured nucleoli. Less cytoplasm is present. |
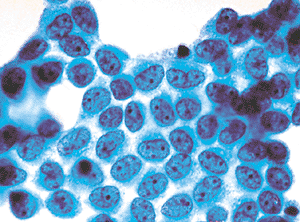 |
Figure 21.9 In this more reactive sheet of mesothelial cells, the cytoplasm has grown smaller and the nuclei relatively larger. Irregularity of the chromatin pattern is more prominent. |
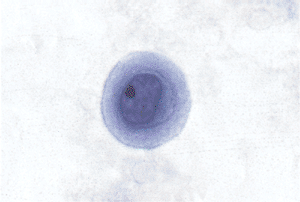 |
Figure 21.10 This individual reactive mesothelial cell has a restricted amount of cytoplasm and a relatively large nucleus with a nucleolus. The cell border is highly irregular and quezzy, suggesting the presence of the numerous elongated microvilli, which are evident on electron microscopy (see Figure 21.24). The cytoplasm is divided into an inner denser layer and an outer less dense layer. Ultrastructurally, the inner more dense layer corresponds to the presence of intermediate filaments with the characteristics of keratin (see Figure 21.25). |
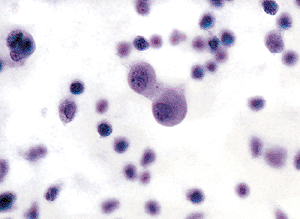 |
Figure 21.11 Mesothelial reaction is frequently associated with inflammatory cells. These reactive mesothelial cells are joined in pairs. Each is several times the size of either neutrophils or lymphocytes. |
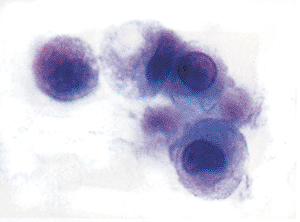 |
Figure 21.12 These reactive mesothelial cells are loosely joined together. The uppermost cell has a vacuole in the cytoplasm, which could be either a vesicle or an intracytoplasmic lumen. |
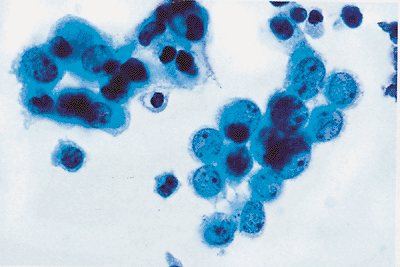 |
Figure 21.13 When reactive mesothelial cells are in groups, an irregular or knobby outside border is formed. Acini form a smooth outer border. |
P.552
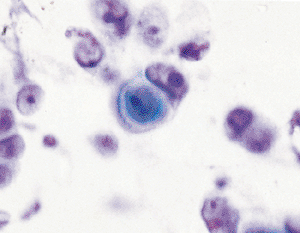 |
Figure 21.14 Occasionally, very reactive mesothelial cells may show cellular interactions similar to that of a keratin pearl. |
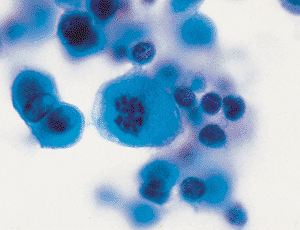 |
Figure 21.15 Mitotic figures may be seen in the proliferating cells of reactive mesothelium. |
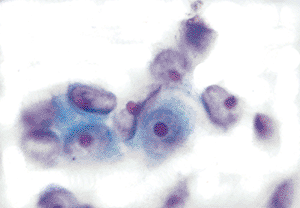 |
Figure 21.16 Markedly reactive mesothelial cells have large vesicular nuclei with a prominent nucleoli. |
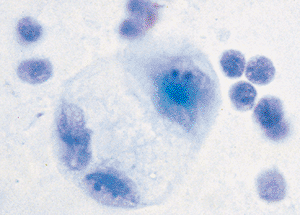 |
Figure 21.17 Reactive mesothelial cells may degenerate and swell. These three cells have abundant multivacuolated cytoplasm and the large nuclei of the reactive mesothelial cells. |
Histochemistry
Histochemical stains have provided information about mucoprotein content of the mesothelium. Negativity for the periodic acid-Schiff (PAS) reaction is evidence that mesothelial cells lack significant quantities of neutral mucoproteins. In contrast, positivity for histochemical stains that detect negative groups, such as the positively charged dye Alcian blue, is evidence of acid mucoproteins. That the intensity of staining reactions for acid mucosubstances is diminished by preincubating the tissue sections in hyaluronidase is evidence that at least some of the terminal hexose groups of the mucosubstances are either hyaluronic acid or chondroitin sulfate (Figure 21.19). Furthermore, the fact that histochemical mucin is decreased, but not abolished, by incubating cells in neuraminidase prior to histochemical
P.553
staining is evidence that some of the terminal carbohydrate groups are sialated (23). MacDougall et al. (24) have documented that neoplastic mesothelial cells may stain with mucicarmine.
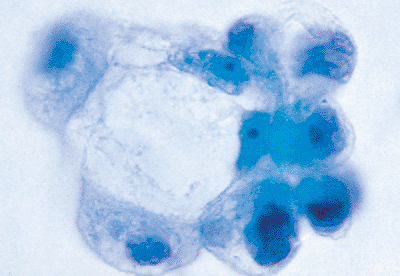 |
Figure 21.18 When markedly reactive mesothelial cells form irregular groups and combine with degenerating forms, they may mimic the appearance of a mucin-producing adenocarcinoma. |
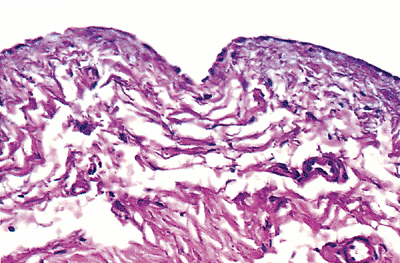 |
Figure 21.19 Alcian blue stain of serosal membrane. Note staining of both mesothelium and of the matrix proteins, which contain abundant acid mucoproteins. |
The types of terminal carbohydrate groups of membrane proteins and lipids can also be characterized with lectins, which have specific and discrete ranges of sugar group affinities. Concanavalin A mesothelial cell reactivity indicates the presence of terminal groups that are either -mannose or -glucose.
Immunohistochemistry
Immunohistochemistry is the most commonly employed method for demonstrating mesothelial differentiation. Mesothelial cells preferentially express a group of so-called mesothelial markers (calretinin, cytokeratin 5/6, WT-1, HBME-1, thrombomodulin, and D2-40) relative to so-called epithelial markers (CEA, CD15, MOC-31, BER-EP4, B72.3, and TTF-1). However, overlap in the expression of any single marker has led to the use of a panel of antibodies to distinguish epithelium and, hence metastatic carcinoma, from reactive mesothelium.
Calretinin, a calcium-binding protein of 29 kDa with similarity to S-100 protein, is found in both the nucleus and cytoplasm of reactive and neoplastic mesothelium but is also found in some adenocarcinomas (25,26,27,28) (Figure 21.20). Cytokeratin 5 is found in the cytoplasm of most mesothelial cells and squamous cell carcinomas, but few adenocarcinomas (29); WT-1, a product of the Wilms' tumor gene, is found in the nucleus of reactive and neoplastic mesothelium and in ovarian surface epithelium and tumors derived therefrom (30). Thrombomodulin, a transmembrane glycoprotein, gives a membranous stain in about half the mesotheliomas but also in some adenocarcinomas. Mesothelial cells also express mesothelin, epithelial membrane antigen (EMA), N-cadherin, E-cadherin, and vimentin (31). Recently, D2-40 (a marker of lymphatic epithelium) has been reported to identify mesothelial cells with a high sensitivity, but it also marks ovarian serous carcinoma (32).
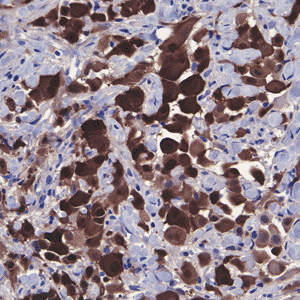 |
Figure 21.20 Immunohistochemical staining of mesothelial cells for calretinin. Note the nuclear and cytoplasmic staining. |
Mesothelial cells usually lack the glycoproteins detected by antibodies to CEA, MOC-31 and BER-EP4 and the determinant detected by Leu-M1 (CD15) (33,34,35,36). Latza et al. (37) and Sheibani et al. (38) reported that BER-EP4 was used to distinguish malignant epithelium (adenocarcinoma) from malignant mesothelioma, but Gaffey et al. (39) and Otis (40) reported BER-EP4 immunoreactivity in high proportions of both benign and malignant mesothelial tumors, as well as adenocarcinomas. The tissue specific nuclear transcription protein TTF-1 is important in embryogenesis of thyroid and lung and is found in nuclei of pneumocytes and many adenocarcinomas of the lung but not in mesothelium (41).
Overlap between reactive and neoplastic mesothelium in the expression of even a panel of antibodies leaves only the demonstration of invasion of parietes or organs to confirm the diagnosis of malignant mesothelioma in most cases. The diagnosis of mesothelioma-in-situ requires demonstration of invasive mesothelioma elsewhere in the same specimen or in a subsequent specimen.
Nonproliferating mesothelial cells express both vimentin and a variety of keratins (K7 of 55 kDa, K8 of 53 kDa, K18 of 44 kDa, and K19 of 40 kDa) that can be detected with monoclonal antibodies immunoreactive with the small, acidic, type I keratins (42) (Figures 21.21,21.22).
P.554
Mesothelium does not express cytokeratin 20, using monoclonal antibodies (43). These keratins are distinct from those of epithelia, including the epidermis, glandular epithelia, and transitional epithelium (44). Ovarian epithelial tumors express a spectrum of keratins similar to that of mesothelium (44).
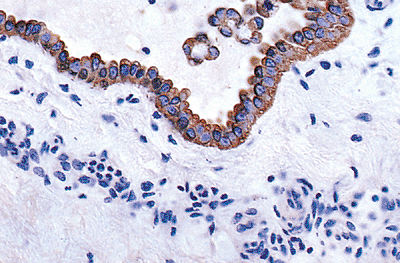 |
Figure 21.21 Keratin expression by mesothelium and detached mesothelial cells, demonstrated by a mixture of monoclonal antikeratin antibodies (AE1/AE3). Note that the immunohistochemical pattern of staining is similar to the distribution of intermediate filaments seen at the ultrastructural level. |
The plasticity of the immunophenotype of mesothelial cells is demonstrable in abnormal states. Although mesothelium normally lacks sex steroid receptors, reactive mesothelium adjacent to endometriosis expresses focal immunoreactivity for estrogen and progesterone receptors (45). Furthermore, reactive mesothelial cells can express the muscle cell cytoskeleton proteins desmin and muscle-specific actin (46). There is experimental evidence that the pattern of intermediate filament expression by mesothelial cells is dependent on shape and cell-cell interaction. Induction of a spindle morphology inhibits keratin synthesis. In contrast, induction of an epithelioid morphology (for example, with retinoids) stimulates keratin synthesis and inhibits vimentin synthesis; the ability of cells to respond in this manner also depends on the presence of cell-cell interactions (47). The pattern of intermediate filament expression by intact mesothelium has not been analyzed in such detail (Figures 21.20,21.21).
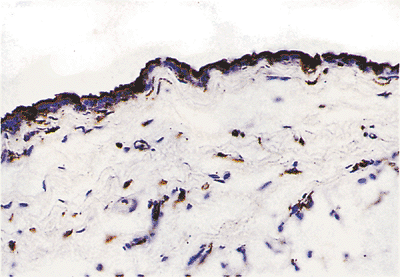 |
Figure 21.22 Vimentin immunoreactivity of mesothelial cells and fibroblasts. |
Ultrastructure
Numerous long microvilli (Figures 21.23,21.24), measuring up to 3 m in length and 0.1 m in diameter, are present and are more numerous in caudal portions of the parietal pleura and in the visceral pleura. The other organelles found in mesothelial cells are not specific for them. Junctions of all types are found tight junctions that serve as a barrier to certain molecules, gap junctions for cell-cell transport, and desmosomes for cell-cell adherence. Intermediate filaments are somewhat prominent; although they do not aggregate into bundles, they are often arranged in a perinuclear, circumferential distribution (Figures 21.25,21.26).
Submesothelial Layer
Normally, the submesothelial layer contains few cells, and most of these are fibroblasts. Much of the submesothelial layer is composed of collagen, elastin, and other extracellular proteins. During reactive processes, the submesothelial layer may become much more prominent as cells proliferate there.
Histochemistry
The main constituents of the submesothelial tissue are glycosylated proteins, including glycosaminoglycans. Because the majority of carbohydrate groups are negatively charged (as a result of an abundance of hyaluronic acid and other acidic groups), this extracellular matrix stains in a manner characteristic of acidic mucoproteins; that is, it is Alcian blue positive (Figure 21.19). That staining intensity can be diminished by treating the section with hyaluronidase before histochemical staining is evidence that hyaluronic acid groups are responsible, in large part, for the intensity of staining (21).
Immunohistochemistry
The antigens of the submesothelial layer can be categorized into matrix constituents and antigens of the mesenchymal cells. The extracellular matrix materials are those typical of most connective tissue. Types I and III collagen and fibronectin are abundant. Elastin fibers are plentiful and basement membrane proteins, including type IV collagen and laminin, are found at the mesothelial cell-stromal interface.
P.555
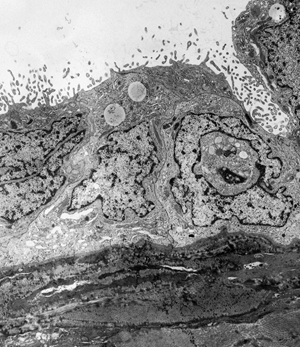 |
Figure 21.23 Mesothelial cells with their elongated microvilli, cover the surface of the serosa. The subjacent stroma is composed of collagen and fibroblasts. |
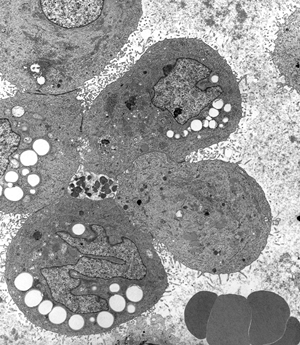 |
Figure 21.24 EM of a cluster of detached mesothelial cells within a pleural effusion. Cytoplasmic lipid droplets impart a vacuolated appearance to some cells. Note the long microvilli, which impart the fuzzy appearance to these cells at the light microscopic level. |
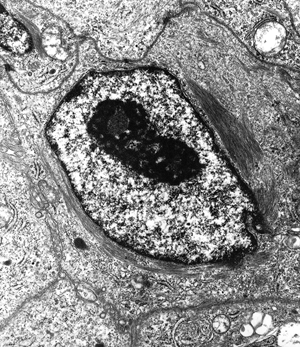 |
Figure 21.25 Ultrastructure of a mesothelial cell. Intermediate filaments are arranged in a perinuclear distribution. |
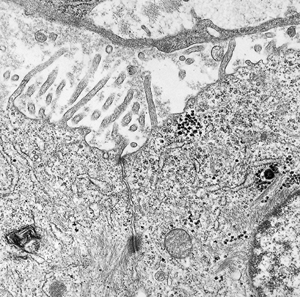 |
Figure 21.26 High magnification of the luminal aspect of two mesothelial cells. Note the small tight junction, subjacent desmosome, and the cytoskeletal filaments within the microvilli. |
P.556
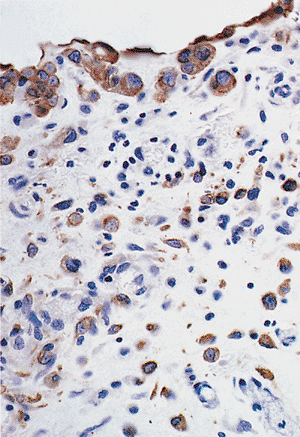 |
Figure 21.27 Keratin (AE1/AE3) immunoreactivity of proliferating submesothelial spindle cells. |
Proteoglycans are plentiful. The pattern of intermediate filament expression by the submesothelial stromal cells varies with their state of excitation. Quiescent cells contain only vimentin, but stromal cells in regions of injury or inflammation also synthesize keratin detectable with antibodies to type I keratins (47) (Figure 21.27).
Relationship of Mesothelial and Submesothelial Cells
The submesothelial mesenchymal cell population serves as the anchoring substratum for the mesothelium. Both mesothelial and submesothelial cells contribute to the extracellular proteins that comprise the matrix. Up to 3% of the total protein synthesized by mesothelial cells are collagens and laminin.
A controversial topic is whether submesothelial cells serve as a source of mesothelial cell renewal, either in normal development, which is the basis for regarding mesothelial cells as mesodermal in origin, or in conditions of rapid mesothelial cell turnover. Earlier ultrastructural and kinetics studies, using thymidine incorporation, suggested that the stromal cells contribute to the repopulation of denuded mesothelium (48,49). Consistent with this scheme is the observation that submesothelial cells, when stimulated to proliferate, synthesize keratin and assume a more epithelioid morphology. However, later studies have demonstrated that healing of injured serosa progresses by multiplication and migration of mesothelial cells at the edges of the wounded area (50).
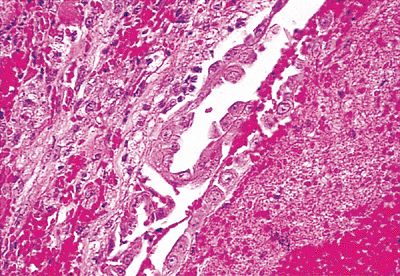 |
Figure 21.28 In a variety of reactive processes, the mesothelial cells lining serosal surfaces undergo markedly proliferative and hyperplastic changes. This is a section through the pleura of a patient with severe active rheumatoid arthritis. |
Reactive Mesothelium
The capacity for mesothelial and submesothelial cellular elements of serous membranes to react and proliferate to produce morphologic patterns mimicking neoplasia is well known and frequently a source of diagnostic confusion (51,52). The process may be diffuse or localized (Figure 21.28). Mesothelial hyperplasia in herniorrhaphy specimens is well described (53) and may be nodular, demonstrate nuclear atypia and frequent mitotic figures, and be accompanied by spindle cell elements (Figures 21.29,21.30).
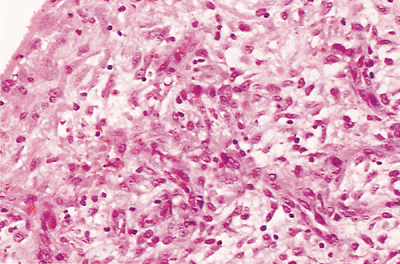 |
Figure 21.29 This photomicrograph is from a hernia sac of an 18-month-old boy. The reactive mesothelium is composed of proliferative epithelioid cells on the surface and subjacent spindle-shaped cells that give the impression of proliferating fibroblasts. |
P.557
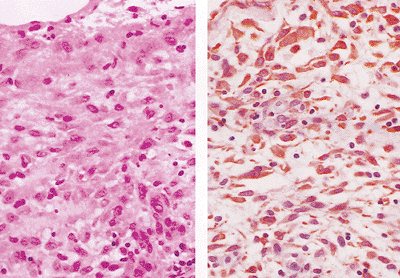 |
Figure 21.30 The reactive peritoneum in the hernia sac shown in Figure 21.29 (A). The immunohistochemical stain for keratin (AE1/AE3) (B) illustrates that both the surface lining mesothelial cells and the subjacent spindle-shaped cells are heavily decorated. |
Reactive mesothelial cells in mediastinal lymph nodes have been reported by Brooks et al. (54) and Parkash et al. (55), although the mechanisms by which they enter lymphatics and survive in the sinuses of nodes are not known. This rare event may produce a difficult differential diagnosis. Clear demonstration of the mesothelial nature of these cells is necessary to exclude metastatic carcinoma. If the cells are shown to be mesothelial by electron microscopy or immunohistochemistry, it should be noted that Sussman and Rosai (56) reported that mesothelioma may present as a lymph node metastasis. Therefore, follow-up may be required to make the distinction between reactive benign mesothelial cells and metastatic malignant ones (Figures 21.31,21.32).
 |
Figure 21.31 Internal mammary lymph node with large epithelioid cells in the sinuses found during coronary artery bypass graft surgery in a 61-year-old man. No pleural lesion was present. |
An uncommon manifestation of mesothelial proliferation is the psammoma body a laminated calcific structure that most likely arises through concentric calcification following cell death. Psammoma bodies are usually nonspecific because they may be observed in inflammatory processes accompanied by mesothelial hyperplasia, malignant mesothelial neoplasia, or epithelial neoplasia (Figure 21.33).
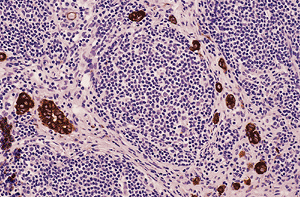 |
Figure 21.32 Immunohistochemical stain for keratin AE1/AE3 demonstrates the epithelioid cells that were negative for CEA, Leu-M1, BER-EP4, and B72.3 and hence are considered reactive mesothelial cells. |
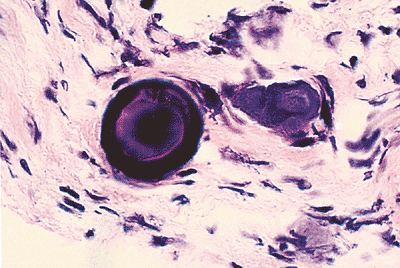 |
Figure 21.33 Psammoma bodies in the left pelvic peritoneum of a 58-year-old female. |
P.558
Endosalpingiosis and Endometriosis
Epithelial elements may be observed in glandular arrangements throughout the peritoneum, omentum, and within lymph nodes. Such glandular structures were recognized in the early 1900s and misinterpreted by some as metastatic carcinoma, a mistake that is unfortunately still committed today. Endometriosis and endosalpingiosis were expounded upon by Sampson (57,58,59) earlier in this century, with reference to mechanisms of pathogenesis that are still debatable.
Endosalpingiosis refers to glandular spaces lined by epithelium similar to uterine tube epithelium, with three cell types (ciliated, secretory, and intercalated cells) (60) (Figure 21.34). On occasion, psammoma bodies are present. Periglandular stroma containing chronic inflammatory cells is separated from epithelium by PAS-positive basement membrane. Endosalpingiosis may be differentiated from endometriosis by the lack of endometrial stroma or evidence of stromal hemorrhage associated with endometriosis (57,60,61,62,63). This condition is seen exclusively in women and has been reported in 12.5% of omenta removed at surgery in females. A large proportion of these women have coexisting benign disease of the uterine tube (60). The origin of the glandular inclusions is debated but is most likely either related to the influence of m llerian development on the peritoneal mesothelium (coelomic lining) or is a sequela of disease within the uterine tube resulting in extratubal growth of displaced tubal epithelium (60,62). Although definitive evidence of neoplasia arising in endosalpingiosis has not been documented (64), considerable difficulty may be encountered when differentiating extraovarian tumor implants removed in the setting of common epithelial ovarian tumors from endosalpingiosis with cellular atypia. Evaluation of the severity of epithelial atypia, mitotic activity, the presence of ciliated cells, and the presence of invasive characteristics may aid in establishing malignancy in this setting (60). Metaplasia in endosalpingiosis may also be a source of diagnostic difficulty particularly mucinous metaplasia, which may be mistaken for metastatic mucinous adenocarcinoma (Figures 21.35,21.36).
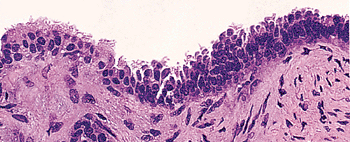 |
Figure 21.34 Endosalpingiosis involving the serosa of the uterus of a 56-year-old woman. Serous, intercalated, and occasional ciliated cells are present, but endometrial stroma is not. |
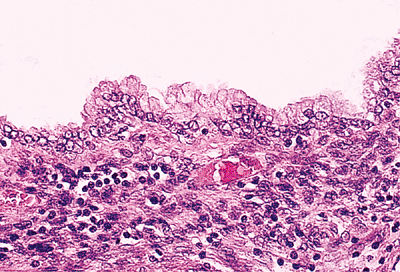 |
Figure 21.35 Endosalpingiosis involving the omentum contains cystic glands lined by mucinous epithelium with basally oriented nuclei and apical cytoplasm. Periglandular stroma contains mononuclear inflammatory cells. |
Endometriosis may be defined by the presence of glands lined by endometrial-type epithelium surrounded by endometrial stroma, outside the uterine endometrial mucosa
P.559
and myometrium (64). The condition occurs most frequently in women of childbearing age. It may occur in a variety of body sites, ranging from the pelvic peritoneum to distant organs such as lung, kidney, and skin, but the most frequent site is the peritoneal lining of the pelvic organs (Figure 21.37). Although the histogenesis of endometriosis remains unclear, two general theories have been proposed. The ectopic growth of endometrial elements may result from displacement of endometrial tissue, whether through local means (such as entry of endometrium into the pelvis through the uterine tubes) or via vascular routes to distant organs (57,58). Another possibility includes metaplastic change of the pelvic peritoneum along m llerian lines of differentiation (65,66). Each mechanism may play a role in the histogenesis of endometriosis.
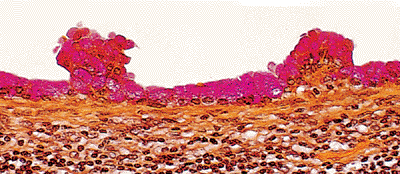 |
Figure 21.36 Mucicarmine stain of mucinous change in endosalpingiosis demonstrates intracytoplasmic mucin in apical cytoplasm. |
Endometriosis may appear as brown-maroon foci on the peritoneal surfaces and be accompanied by fibrosis or adhesions. Microscopically, endometrial stroma surrounding endometrial epithelium is present (67). Response to hormonal influences is often seen and may be synchronous with intrauterine endometrium. Metaplasia occurs in both epithelial and stromal elements, similar to metaplasias encountered in the endometrium of the uterus. The presence of hemosiderin-laden macrophages and fibrosis may be the only evidence that endometriosis had once been present. However, a definitive diagnosis of endometriosis may not be rendered unless both endometrial glands and stroma are seen.
Another common type of metaplasia, more frequently observed in pregnant than in nonpregnant women, is decidual change. Although usually encountered in the submesothelial layer of pelvic peritoneal surfaces, decidual change may be seen in distant sites including the serosal surfaces of the liver, spleen, diaphragm, and within lymph nodes. In these locations, decidual change may be mistaken for metastatic carcinoma or malignant mesothelioma (Figure 21.38) (66).
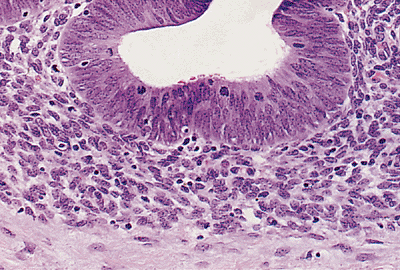 |
Figure 21.37 Endometriosis involving the peritoneum with extension into the soft tissue of the anterior abdominal wall of a 23-year-old woman. Endometrial glands and stroma are present. |
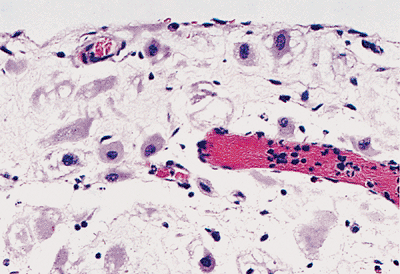 |
Figure 21.38 Decidual change in the pelvis during pregnancy is seen in subserosal tissue. Loosely cohesive cells with abundant eosinophilic cytoplasm are present. |
Fibrous Pleurisy
Fibrous pleurisy is a benign reactive process that usually occurs in the setting of organizing pleural effusions. The differential diagnosis of fibrous pleurisy and desmoplastic mesothelioma may be extremely difficult: both may have regions of increased cellularity in a predominantly fibrous background containing spindle cells that are immunoreactive to keratin antibodies. Fibrous pleurisy tends to have a higher cellularity immediately beneath the fibrinous exudative surface of the pleura and demonstrates a layering of spindle cells parallel to the fibrosis with intervening fibrinous exudate. This organization imparts a histologic sense of order to the reactive process that may assist in its recognition. Invasion, bland necrosis, and sarcomatous foci are not seen in fibrous pleurisy (67,68) (Figure 21.39).
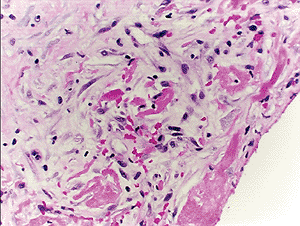 |
Figure 21.39 The histologic appearance of fibrous pleurisy reflects its inflammatory nature, with granulation tissue, fibrin, and a zonal pattern ranging from active inflammation to quiescent dense fibrosis. |
P.560
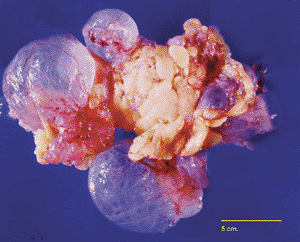 |
Figure 21.40 Multilocular peritoneal inclusion cyst in the omentum of a 73-year-old patient discovered incidentally at surgery for urogynecologic repair procedure. The cysts vary in size, some being translucent while others are fibrotic, particularly toward the center of the mass. |
Multilocular Peritoneal Inclusions Cyst
Multilocular peritoneal inclusions cyst (MPIC) is a mesothelial-lined multilocular lesion that occurs almost exclusively in women. The lesion usually involves the pelvis, although it may occur in other abdominal locations, including the omentum and mesentery. Usually MPIC is mass forming and may attain diameters up to 20 cm. Grossly, it is composed of multiple cysts, some of which may be thinwalled and translucent (Figure 21.40). Histologically, the septa range from thin and delicate to thickened and inflamed. The mesothelial lining ranges from single flattened cells to hobnail-type cells. Squamous metaplasia of the lining mesothelium may be present. Some regions may resemble the cellular pattern of an adenomatoid tumor (Figures 21.41,21.42) (69).
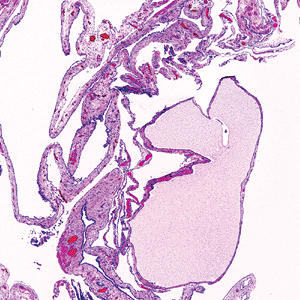 |
Figure 21.41 Histologically, multilocular peritoneal inclusions cysts (MPIC) reflect the gross features, with septae that vary in thickness and cysts that vary in size. |
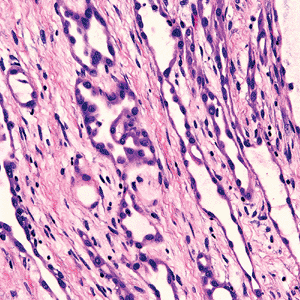 |
Figure 21.42 Some regions in multilocular peritoneal inclusions cysts may contain mesothelial proliferations that closely resemble an adenomatoid tumor. |
The true nature of MPIC remains somewhat controversial, with some authors maintaining that it is a neoplasm while others assert it is a reactive lesion that develops in response to injury or even endometriosis. The original designation of multicystic mesothelioma reflects the notion that the lesion is neoplastic. Recurrences are frequent, although MPIC-related deaths probably do not occur (70).
References
1. Sahn SA. State of the art: the pleura. Am Rev Respir Dis 1988;138:184 234.
2. Pistolesi M, Miniati M, Giuntini C. Pleural liquid and solute exchanges. Am Rev Respir Dis 1989;140:825 847.
3. Courtice FC, Simmonds WJ. Absorption of fluids from the pleural cavities of rabbits and cats. J Physiol 1949;109:117 130.
4. Albertine KH, Wiener-Kronish JP, Roos PJ, Staub NC. Structure, blood supply, and lymphatic vessels of the sheep's visceral pleura. Am J Anat 1982;:165:227 294.
5. Grant T, Levin B. Lymphangiographic visualization of pleural and pulmonary lymphatics in a patient without a chylothorax. Radiology 1974;113:49 50.
P.561
6. Gallagher B, Urbanski SJ. The significance of pleural elastica invasion by lung carcinomas. Hum Pathol 1990;21:512 517.
7. Mariassy AT, Wheeldon EB. The pleura: a combined light microscopic, scanning, and transmission electron microscopic study in the sheep. I. Normal pleura. Exp Lung Res 1983;4:293 314.
8. Albertine KM, Wiener-Kronish JP, Staub NC. The structure of the parietal pleura and its relationship to pleural liquid dynamics in sheep. Anat Rec 1984;208:401 409.
9. Leak LV. Gross and ultrastructural morphologic features of the diaphragm. Am Rev Respir Dis 1979;119(pt 2):3 21.
10. Wang NS. The preformed stomas connecting the pleural cavity and the lymphatics in the parietal pleura. Am Rev Respir Dis 1975;111:12 20.
11. Wang NS. Morphological data of pleura. Normal conditions. In: Chretien J, Hirsch A, eds. Diseases of the Pleura. New York: Masson; 1983:10 24.
12. Wang NS. Anatomy and physiology of the pleural space. Clin Chest Med 1985;6:3 16.
13. Bernaudin JF, Fleury J. Anatomy of the blood and lymphatic circulation of the pleural serosa. In: Chretien J, Bignon J, Hirsch A, eds. The Pleura in Health and Disease. Vol. 30. New York: Marcel Dekker; 1985:101 124.
14. Cooray GH. Defense mechanism in the mediastinum with special reference to the mechanism of pleural absorptions. J Pathol Bacteriol 1949;6:551 567.
15. Courtice FC, Simmonds WJ. Physiological significance of lymph drainage of the serous cavities and lungs. Physiol Rev 1954;34:419 448.
16. Staub NC, Wiener-Kronish JP, Albertine KH. Transport through the pleura. Physiology of normal liquid and solute exchange in the pleural space. In: Chretien J, Bignon J, Hirsch A, eds. The Pleura in Health and Disease. New York: Marcel Dekker; 1985:169 193.
17. Kampmeier OF. Concerning certain mesothelial thickenings and vascular plexuses of the mediastinal pleura associated with histiocyte and fat-cell production, in the human newborn. Anat Rec 1928;39:201 208.
18. Mixter RL. On macrophagal foci ( milky spots ) in the pleura of different mammals, including man. Am J Anat 1941;69:159 186.
19. Nagel W, Kuschinsky W. Study of the permeability of the isolated dog mesentery. Eur J Clin Invest 1970;1:149 154.
20. Tslibari E, Wissig SL. Lymphatic absorption from the peritoneal cavity: regulation of patency of mesothelioma stomata. Microvasc Res 1983;25:220 239.
21. Chretien J, Bignon J, Hirsch A, eds. The Pleura in Health and Disease. Vol 30. New York: Marcel Dekker; 1985.
22. Ernst CS, Brooks JJ. Immunoperoxidase localization of secretory component in reactive mesothelium and mesotheliomas. J Histochem Cytochem 1981;29:1102 1104.
23. Roth J. Ultrahistochemical demonstration of saccharide components of complex carbohydrates at the alveolar cell surface and at the mesothelial cell surface of the pleura visceralis of mice by means of concanavalin A. Exp Pathol (Jena) 1973;8:157 167.
24. MacDougall DB, Wang SE, Zidar BL. Mucin-positive epithelial mesothelioma. Arch Pathol Lab Med 1992;116:874 880.
25. Doglioni C, Tos AP, Laurino L, et al. Calretinin: a novel immunocytochemical marker for mesothelioma. Am J Surg Path 1996;20:1037 1046.
26. Nagel H, Hemmerlein B, Ruschenburg I, Huppe K, Droese M. The value of anti-calretinin antibody in the differential diagnosis of normal and reactive mesothelia versus metastatic tumors in effusion cytology. Pathol Res Pract 1998;194:759 764.
27. Oates J, Edwards C. HBME-1, MOC-31, WT1 and calretinin: an assessment of recently described markers for mesothelioma and adenocarcinoma. Histopathology 2000;36:341 347.
28. Fetsch PA, Simsir A, Abati A. Comparison of antibodies to HBME-1 and calretinin for the detection of mesothelial cells in effusion cytology. Diagn Cytopahtol 2001;25:158 161.
29. Chu PG, Weiss LM. Expression of cytokeratin 5/6 in epithelial neoplasms: an immunohistochemical study of 509 cases. Mod Pathol 2002;15:6 10.
30. Hecht JL, Lee BH, Pinkus JL, Pinkus GS. The value of Wilms tumor susceptibility gene 1 in cytologic preparations as a marker for malignant mesothelioma. Cancer 2002;96:105 109.
31. Ordonez NG. The immunohistochemical diagnosis of mesothelioma: a comparative study of epithelioid mesothelioma and lung adenocarcinoma. Am J Surg Pathol 2003;27:1031 1051.
32. Chu AY, Litzky LA, Pasha TL, Acs G, Zhang PJ. Utility of D2-40, a novel mesothelial marker, in the diagnosis of malignant mesothelioma. Mod Pathol 2005;18:105 110.
33. Otis CN, Carter D, Cole S, Battifora H. Immunohistochemical evaluation of pleural mesothelioma and pulmonary adenocarcinoma. A bi-institutional study of 47 cases. Am J Surg Pathol 1987;11:445 456.
34. Sheibani K, Battifora H, Burke JS, Rappaport H. Leu-M1 antigen in human neoplasms: an immunohistologic study of 400 cases. Am J Surg Pathol 1986;10:227 236.
35. Sheibani K, Esteban JM, Bailey A, Battifora H, Weiss LM. Immunopathologic and molecular studies as an aid to the diagnosis of malignant mesothelioma. Hum Pathol 1992;23:107 116.
36. Sheibani K. Immunopathology of malignant mesothelioma. Hum Pathol 1994;25:219 220.
37. Latza U, Niedobitek G, Schwarting R, Nekarda H, Stein H. Ber-EP4: new monoclonal antibody which distinguishes epithelia from mesothelia. J Clin Pathol 1990;43:213 219.
38. Sheibani K, Shin SS, Kezirian J, Weiss LM. Ber-EP4 antibody as a discriminant in the differential diagnosis of malignant mesothelioma versus adenocarcinoma. Am J Surg Pathol 1991;15:779 784.
39. Gaffey MJ, Mills SE, Swanson PE, Zarbo RJ, Shah AR, Wick MR. Immunoreactivity for Ber-EP4 in adenocarcinomas, adenomatoid tumors, and malignant mesotheliomas. Am J Surg Pathol 1992;16:593 599.
40. Otis CN. Uterine adenomatoid tumors: immunohistochemical characteristics with emphasis on Ber-EP4 immunoreactivity and distinction from adenocarcinoma. Int J Gynecol Pathol 1996;15:146 151.
41. Ordonez NG. Value of thyroid transcription factor-1, E-cadherin, BG8, WT1 and CD44S immunostaining in distinguishing epithelial pleural mesothelioma from pulmonary and nonpulmonary adenocarcinoma. Am J Surg Pathol 2000;24:598 606.
42. Wu YJ, Parker LM, Binder NE, et al. The mesothelial keratins: a new family of cytoskeletal proteins identified in cultured mesothelial cells and nonkeratinizing epithelia. Cell 1982;31(pt 2):693 703.
43. Moll R, Lowe A, Laufer J, Franke WW. Cytokeratin 20 in human carcinomas. A new histodiagnostic marker detected by monoclonal antibodies. Am J Pathol 1992;140:427 447.
44. Moll R, Franke WW, Schiller DL, Geiger B, Krepler R. The catalog of human cytokeratins: patterns of expression in normal epithelia, tumors and cultured cells. Cell 1982;31:11 24.
45. Nakayama K, Masuzawa H, Li S, et al. Immunohistochemical analysis of the peritoneum adjacent to endometriotic lesions using antibodies for Ber-EP4 antigen, estrogen receptors, and progesterone receptors: implication of peritoneal metaplasia in the pathogenesis of endometriosis. Int J Gynecol Pathol 1994;13:348 358.
46. Pitt MA, Haboubi NY. Serosal reaction in chronic gastric ulcers: an immunohistochemical and ultrastructural study. J Clin Pathol 1995;48:226 228.
47. Bolen JW, Hammer SP, McNutt MA. Reactive and neoplastic serosal tissue. A light-microscopic, ultrastructural and immunocytochemical study. Am J Surg Path 1986;10:34 47.
48. Raftery AT. Regeneration of parietal and visceral peritoneum in the immature animal: a light and electron microscopical study. Br J Surg 1973;60:969 975.
49. Raftery AT. Regeneration of parietal and visceral peritoneum: an electron microscopical study. J Anat 1984;115(pt 3):375 392.
50. Whitaker D, Papadimitriou JM. Mesothelial healing: morphological and kinetic investigations. J Pathol 1985;145:159 175.
51. Ackerman LV. Tumors of the retroperitoneum, mesentery, and peritoneum. In: Atlas of Tumor Pathology. 6th series, fascicle 23, 24. Washington, DC; Armed Forces Institute of Pathology; 1954.
52. McCaughey WTE, Kannerstein M, Churg J. Tumors and pseudotumors of the serous membranes. In: Atlas of Tumor Pathology. 2nd series, fascicle 20. Washington, DC: Armed Forces Institute of Pathology; 1983.
P.562
53. Rosai J, Dehner LP. Nodular mesothelial hyperplasia in hernia sacs: a benign reactive condition simulating a neoplastic process. Cancer 1975;35:165 175.
54. Brooks JS, LiVolsi VA, Pietra GG. Mesothelial cell inclusions in mediastinal lymph nodes mimicking metastatic carcinoma. Am J Clin Pathol 1990;93:741 748.
55. Parkash V, Vidwans M, Carter D. Benign mesothelial cells in mediastinal lymph nodes. Am J Surg Pathol 1999;23:1264 1269.
56. Sussman J, Rosai J. Lymph node metastasis as the initial manifestation of malignant mesothelioma: report of six cases. Am J Surg Path 1990;14:819 828.
57. Sampson JA. Heterotopic or misplaced endometrial tissue. Am J Obstet Gynecol 1925;10:649 664.
58. Sampson JA. Postsalpingectomy endometriosis (endosalpingiosis). Am J Obstet Gynecol 1930;20:443 480.
59. Sampson JA. The pathogenesis of postsalpingectomy endometriosis in laparotomy scars. Am J Obstet Gynecol 1946;50:597 620.
60. Zinsser KR, Wheeler JE. Endosalpingiosis in the omentum: a study of autopsy and surgical material. Am J Surg Pathol 1982;6:109 117.
61. Hsu YK, Parmley TH, Rosenshein NB, Bhagavan BS, Woodruff JD. Neoplastic and non-neoplastic mesothelial proliferations in pelvic lymph nodes. Obstet Gynecol 1980;55:83 88.
62. Horn LC, Bilek K. Frequency and histogenesis of pelvic retroperitoneal lymph node inclusions of the female genital tract. An immunohistochemical study of 34 cases. Pathol Res Pract 1995;191:991 996.
63. Schnurr RC, Delgado G, Chun B. Benign glandular inclusions in para-aortic lymph nodes in women undergoing lymphadenectomies. Am J Obstet Gynecol 1978;130:813 816.
64. Clement PB. Endometriosis, lesions of the secondary m llerian system, and pelvic mesothelial proliferations. In: Kurman RJ, ed. Blaustein's Pathology of the Female Genital Tract. 3rd ed. New York: Springer-Verlag;1987:517 559.
65. Ferguson BR, Bennington JL, Haber SL. Histochemistry of mucosubstances and histology of mixed m llerian pelvic lymph node glandular inclusions: evidence for histogenesis by m llerian metaplasia on coelomic epithelium. Obstet Gynecol 1969;33:617 625.
66. Lauchlan SC. The secondary m llerian system. Obstet Gynecol Surv 1972;27:133 146.
67. Churg A, Colby TV, Cagle P, et al. The separation of benign and malignant mesothelial proliferations. Am J Surg Pathol 2000;24:1183 1200.
68. Mangano WE, Cagle PT, Churg A, Vollmer RT, Roggli VL. The diagnosis of desmoplastic malignant mesothelioma and its distinction from fibrous pleurisy: a histologic and immunohistochemical analysis of 31 cases including p53 immunostaining. Am J Clin Pathol 1998;110:191 199.
69. Clement PB. Reactive tumor-like lesions of the peritoneum. Am J Clin Pathol 1995;103:673 676.
70. Weiss SW, Tavassoli FA. Multicystic mesothelioma. An analysis of pathologic findings and biologic behavior in 37 cases. Am J Surg Pathol 1988;12:737 746.