 | Note: Large images and tables on this page may necessitate printing in landscape mode.
Copyright 2006 The McGraw-Hill Companies. All rights reserved.
Lange Cardiovascular Physiology > Chapter 1. Overview of the Cardiovascular System >
| Objectives The student understands the homeostatic role of the cardiovascular system, the basic principles of cardiovascular transport, and the basic structure and function of the components of the system. - Defines homeostasis.
- Identifies the major body fluid compartments and states the approximate volume of each.
- Lists two conditions, provided by the cardiovascular system that are essential for regulating the composition of interstitial fluid (ie, the internal environment).
- Diagrams the blood flow pathways between the heart and other major body organs.
- States the relationship among blood flow, blood pressure, and vascular resistance.
- Predicts the relative changes in flow through a tube caused by changes in tube length, tube radius, fluid viscosity, and pressure difference.
- Identifies the chambers and valves of the heart and describes the pathway of blood flow through the heart.
- Defines cardiac output.
- Describes the pathway of action potential propagation in the heart.
- Lists the five factors essential to proper ventricular pumping action.
- States the relationship between ventricular filling and cardiac output (Starling's Law of the Heart) and describes its importance in the control of cardiac output.
- Identifies the distribution of sympathetic and parasympathetic nerves in the heart and lists the basic effects of these nerves on the heart.
- Lists the major different types of vessels in a vascular bed and describes the morphological differences among them.
- Describes the basic anatomical features and function of the different vessel types.
- Identifies the major mechanisms in vascular control and blood flow distribution.
- Describes the basic composition of the fluid and cellular portions of blood.
| | Homeostatic Role of the Cardiovascular System A nineteenth-century French physiologist, Claude Bernard (1813 1878), first recognized that all higher organisms actively and constantly strive to prevent the external environment from upsetting the conditions necessary for life within the organism. Thus the temperature, oxygen concentration, pH, ionic composition, osmolarity, and many other important variables of our internal environment are closely controlled. This process of maintaining the "constancy" of our internal environment has come to be known as homeostasis. To accomplish this task, an elaborate material transport network, the cardiovascular system, has evolved. Three compartments of watery fluids, known collectively as the total body water, account for about 60% of body weight. This water is distributed among the intracellular, interstitial, and plasma compartments, as indicated in Figure 1 1. Note that about two thirds of our body water is contained within cells and communicates with the interstitial fluid across the plasma membranes of cells. Of the fluid that is outside cells (ie, extracellular fluid), only a small amount, the plasma volume, circulates within the cardiovascular system. Blood is composed of plasma and roughly an equal volume of formed elements (primarily red cells). The circulating plasma fluid communicates with the interstitial fluid across the walls of small capillary vessels within organs.
The interstitial fluid is the immediate environment of individual cells. (It is the "internal environment" referred to by Bernard.) These cells must draw their nutrients from and release their products into the interstitial fluid. The interstitial fluid cannot, however, be considered a large reservoir for nutrients or a large sink for metabolic products since its volume is less than half that of the cells that it serves.  The well-being of individual cells therefore depends heavily on the homeostatic mechanisms that regulate the composition of the interstitial fluid. This task is accomplished by continuously exposing the interstitial fluid to "fresh" circulating plasma fluid. The well-being of individual cells therefore depends heavily on the homeostatic mechanisms that regulate the composition of the interstitial fluid. This task is accomplished by continuously exposing the interstitial fluid to "fresh" circulating plasma fluid.
As blood passes through capillaries, solutes exchange between plasma and interstitial fluid by the process of diffusion. The net result of transcapillary diffusion is always that interstitial fluid tends to take on the composition of the incoming blood. If, for example, potassium ion concentration in the interstitium of a particular skeletal muscle were higher than that in the plasma entering the muscle, potassium would diffuse into the blood as it passed through the muscle's capillaries. Since this removes potassium from the interstitial fluid, interstitial potassium ion concentration would decrease. It would stop decreasing when net movement of potassium into capillaries no longer occurred, ie, when interstitial concentration reached that of incoming plasma. Two conditions are essential for this circulatory mechanism to effectively control the composition of interstitial fluid: (1) there must be adequate blood flow through the tissue capillaries, and (2) the chemical composition of the incoming (or arterial) blood must be controlled to be that which is optimal in the interstitial fluid. Figure 1 1 shows how the cardiovascular transport system operates to accomplish these tasks. As discussed above, substances are transported between cells and plasma in capillary vessels within organs by the process of diffusion. This transport occurs over extremely small distances because no cell in the body is located farther than about 10  m from a capillary. Over such microscopic distances, diffusion is a very rapid process that can move huge quantities of material. Diffusion, however, is a very poor mechanism by which to move substances from the capillaries of one organ, such as the lungs, to the capillaries of another organ that may be 1 meter or more distant. Consequently, substances are transported between organs by the process of convection, by which the substances move along with blood flow simply because they are dissolved or otherwise contained within blood. The relative distances involved in cardiovascular transport are not well illustrated in Figure 1 1. If the figure were drawn to scale with 1 inch representing the distance from capillaries to cells within a calf muscle, the capillaries in the lungs would have to be located about 15 miles away! m from a capillary. Over such microscopic distances, diffusion is a very rapid process that can move huge quantities of material. Diffusion, however, is a very poor mechanism by which to move substances from the capillaries of one organ, such as the lungs, to the capillaries of another organ that may be 1 meter or more distant. Consequently, substances are transported between organs by the process of convection, by which the substances move along with blood flow simply because they are dissolved or otherwise contained within blood. The relative distances involved in cardiovascular transport are not well illustrated in Figure 1 1. If the figure were drawn to scale with 1 inch representing the distance from capillaries to cells within a calf muscle, the capillaries in the lungs would have to be located about 15 miles away! The overall functional arrangement of the cardiovascular system is illustrated in Figure 1 2. Since a functional rather than an anatomical viewpoint is expressed in this figure, the heart appears in three places: as the right heart pump, as the left heart pump, and as the heart muscle tissue. It is common practice to view the cardiovascular system as (1) the pulmonary circulation, composed of the right heart pump and the lungs, and (2) the systemic circulation, in which the left heart pump supplies blood to the systemic organs (all structures except the gas exchange portion of the lungs). The pulmonary and systemic circulations are arranged in series, ie, one after the other. Consequently, the right and left hearts must each pump an identical volume of blood each minute. This amount is called the cardiac output. A cardiac output of 5 to 6 L/min is normal for a resting individual. As indicated in Figure 1 2, the systemic organs are functionally arranged in parallel (ie, side by side) within the cardiovascular system. There are two important consequences of this parallel arrangement. First, nearly all systemic organs receive blood of identical composition that which has just left the lungs and is known as arterial blood. Second, the flow through any one of the systemic organs can be controlled independently of the flow through the other organs. Thus, for example, the cardiovascular response to whole body exercise can involve increased blood flow through some organs, decreased blood flow through others, and unchanged blood flow through yet others. Many of the organs in the body help perform the task of continually reconditioning the blood circulating in the cardiovascular system. Key roles are played by organs, such as the lungs, that communicate with the external environment. As is evident from the arrangement shown in Figure 1 2, any blood that has just passed through a systemic organ returns to the right heart and is pumped through the lungs, where oxygen and carbon dioxide are exchanged. Thus the blood's gas composition is always reconditioned immediately after leaving a systemic organ. Like the lungs, many of the systemic organs also serve to recondition the composition of blood, although the flow circuitry precludes their doing so each time the blood completes one circuit. The kidneys, for example, continually adjust the electrolyte composition of the blood passing through them. Because the blood conditioned by the kidneys mixes freely with all the circulating blood and because electrolytes and water freely pass through most capillary walls, the kidneys control the electrolyte balance of the entire internal environment. To achieve this, it is necessary that a given unit of blood pass often through the kidneys. In fact, the kidneys (under resting conditions) normally receive about one fifth of the cardiac output. This greatly exceeds the amount of flow that is necessary to supply the nutrient needs of the renal tissue. This situation is common to organs that have a blood-conditioning function. Blood-conditioning organs can also withstand, at least temporarily, severe reductions of blood flow. Skin, for example, can easily tolerate a large reduction in blood flow when it is necessary to conserve body heat. Most of the large abdominal organs also fall into this category. The reason is simply that because of their blood-conditioning functions, their normal blood flow is far in excess of that necessary to maintain their basal metabolic needs. The brain, heart muscle, and skeletal muscles typify organs in which blood flows solely to supply the metabolic needs of the tissue. They do not recondition blood for the benefit of any other organ. Flow to brain and heart muscle is normally only slightly greater than that required for their metabolism, and they do not tolerate blood flow interruptions well. Unconsciousness can occur within a few seconds after stoppage of cerebral flow, and permanent brain damage can occur in as little as 4 minutes without flow. Similarly, the heart muscle (myocardium) normally consumes about 75% of the oxygen supplied to it, and the heart's pumping ability begins to deteriorate within beats of a coronary flow interruption. As we shall see later, the task of providing adequate blood flow to the brain and the heart muscle receives a high priority in the overall operation of the cardiovascular system. | | The Basic Physics of Blood Flow As outlined above, the task of maintaining interstitial homeostasis requires that an adequate quantity of blood flow continuously through each of the millions of capillaries in the body. In a resting individual, this adds up to a cardiac output of about 5 L/min (about 80 gallons/h). As people go about their daily lives, the metabolic rates and therefore the blood flow requirements in different organs and regions throughout the body change from moment to moment. Thus, the cardiovascular system must continuously adjust both the magnitude of cardiac output and how that cardiac output is distributed to different parts of the body. One of the most important keys to comprehending how the cardiovascular system operates is a thorough understanding of the relationship among the physical factors that determine the rate of fluid flow through a tube.  The tube depicted in Figure 1 3 might represent a segment of any vessel in the body. It has a certain length (L) and a certain internal radius (r) through which blood flows. Fluid flows through the tube only when the pressures in the fluid at the inlet and outlet ends (Pi and Po) are unequal, ie, when there is a pressure difference ( The tube depicted in Figure 1 3 might represent a segment of any vessel in the body. It has a certain length (L) and a certain internal radius (r) through which blood flows. Fluid flows through the tube only when the pressures in the fluid at the inlet and outlet ends (Pi and Po) are unequal, ie, when there is a pressure difference ( P) between the ends. Pressure differences supply the driving force for flow. Because friction develops between moving fluid and the stationary walls of a tube, vessels tend to resist fluid movement through them. This vascular resistance is a measure of how difficult it is to make fluid flow through the tube, ie, how much of a pressure difference it takes to cause a certain flow. The all-important relation among flow, pressure difference, and resistance is described by the basic flow equation as follows: P) between the ends. Pressure differences supply the driving force for flow. Because friction develops between moving fluid and the stationary walls of a tube, vessels tend to resist fluid movement through them. This vascular resistance is a measure of how difficult it is to make fluid flow through the tube, ie, how much of a pressure difference it takes to cause a certain flow. The all-important relation among flow, pressure difference, and resistance is described by the basic flow equation as follows:

The basic flow equation may be applied not only to a single tube but to complex networks of tubes, eg, to the vascular bed of an organ or to the entire systemic system. The flow through the brain, for example, is determined by the difference in pressure between cerebral arteries and veins divided by the overall resistance to flow through the vessels in the cerebral vascular bed. It should be evident from the basic flow equation that there are only two ways in which blood flow through any organ can be changed: (1) by changing the pressure difference across its vascular bed, or (2) by changing its vascular resistance. Most often, it is changes in an organ's vascular resistance that cause the flow through the organ to change. From the work of the French physician Jean Leonard Marie Poiseuille (1799 1869), who performed experiments on fluid flow through small glass capillary tubes, it is known that the resistance to flow through a cylindrical tube depends on several factors, including the radius and length of the tube and the viscosity of the fluid flowing through it. These factors influence resistance to flow as follows: 
Note that the internal radius of the tube is raised to the fourth power in this equation. Thus even small changes in the internal radius of a tube have a very large influence on its resistance to flow. For example, halving the inside radius of a tube will increase its resistance to flow by 16-fold. The preceding equations may be combined into one expression known as the Poiseuille equation, which includes all the terms that influence flow through a cylindrical vessel.2 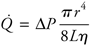
Again note that flow occurs only when a pressure difference exists. It is not surprising then that arterial blood pressure is an extremely important and carefully regulated cardiovascular variable. Also note once again that, for any given pressure difference, tube radius has a very large influence on the flow through a tube. It is logical, therefore, that organ blood flows are regulated primarily through changes in the radius of vessels within organs. Whereas vessel length and blood viscosity are factors that influence vascular resistance, they are not variables that can be easily manipulated for the purpose of moment-to-moment control of blood flow. In regard to the overall cardiovascular system as depicted in Figures 1 1 and 1 2, one can conclude that blood flows through the vessels within an organ only because a pressure difference exists between the blood in the arteries supplying the organ and the veins draining it. The primary job of the heart pump is to keep the pressure within arteries higher than that within veins. Normally, the average pressure in systemic arteries is near 100 mmHg, and the average pressure in systemic veins is near 0 mmHg. Therefore, because the pressure difference (P) is identical across all systemic organs, cardiac output is distributed among the various systemic organs solely on the basis of their individual resistances to flow. Because blood flows along the path of least resistance, organs with relatively low resistance receive relatively high flow. 1 Although pressure is most correctly expressed in units of force per unit area, it is customary to express pressures within the cardiovascular system in millimeters of mercury. For example, mean arterial pressure may be said to be 100 mmHg because it is the same as the pressure existing at the bottom of a mercury column 100 mm high. All cardiovascular pressures are expressed relative to atmospheric pressure, which is approximately 760 mmHg. 2 Poiseuille's equation properly applies only to a homogeneous fluid flowing through rigid nontapered tubes with a certain flow pattern called laminar flow. Although not all these conditions are rigidly met for any vessel within the body, the approximation is close enough to permit general conclusions to be drawn from Poiseuille's equation. | | The Heart Pumping Action The heart lies in the center of the thoracic cavity suspended by its attachments to the great vessels within a thin fibrous sac called the pericardium. A small amount of fluid in the sac lubricates the surface of the heart and allows it to move freely during contraction and relaxation. Blood flow through all organs is passive and occurs only because arterial pressure is kept higher than venous pressure by the pumping action of the heart. The right heart pump provides the energy necessary to move blood through the pulmonary vessels, and the left heart pump provides the energy to move blood through the systemic organs. The amount of blood from each ventricle pumped/minute (the cardiac output, CO) depends on the volume of blood ejected per beat (the stroke volume, SV) and the number of heart beats per minute (the heart rate, HR) as follows: 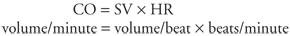
It should be evident from this relationship that all influences on cardiac output must act by changing either the heart rate or the stroke volume. These influences will be described in detail in subsequent chapters. The pathway of blood flow through the chambers of the heart is indicated in Figure 1 4. Venous blood returns from the systemic organs to the right atrium via the superior and inferior venae cavae. It passes through the tricuspid valve into the right ventricle and from there is pumped through the pulmonic valve into the pulmonary circulation via the pulmonary arteries. Oxygenated pulmonary venous blood flows in pulmonary veins to the left atrium and passes through the mitral valve into the left ventricle. From there it is pumped through the aortic valve into the aorta to be distributed to the systemic organs.
 Although the gross anatomy of the right heart pump is somewhat different from that of the left heart pump, the pumping principles are identical. Each pump consists of a ventricle, which is a closed chamber surrounded by a muscular wall, as illustrated in Figure 1 5. The valves are structurally designed to allow flow in only one direction and passively open and close in response to the direction of the pressure differences across them. Ventricular pumping action occurs because the volume of the intraventricular chamber is cyclically changed by rhythmic and synchronized contraction and relaxation of the individual cardiac muscle cells that lie in a circumferential orientation within the ventricular wall. Although the gross anatomy of the right heart pump is somewhat different from that of the left heart pump, the pumping principles are identical. Each pump consists of a ventricle, which is a closed chamber surrounded by a muscular wall, as illustrated in Figure 1 5. The valves are structurally designed to allow flow in only one direction and passively open and close in response to the direction of the pressure differences across them. Ventricular pumping action occurs because the volume of the intraventricular chamber is cyclically changed by rhythmic and synchronized contraction and relaxation of the individual cardiac muscle cells that lie in a circumferential orientation within the ventricular wall.
When the ventricular muscle cells are contracting, they generate a circumferential tension in the ventricular walls that causes the pressure within the chamber to increase. As soon as the ventricular pressure exceeds the pressure in the pulmonary artery (right pump) or aorta (left pump), blood is forced out of the chamber through the outlet valve as shown in Figure 1 5. This phase of the cardiac cycle during which the ventricular muscle cells are contracting is called systole. Because the pressure is higher in the ventricle than in the atrium during systole, the inlet or atrioventricular (AV) valve is closed. When the ventricular muscle cells relax, the pressure in the ventricle falls below that in the atrium, the AV valve opens, and the ventricle refills with blood as shown on the right of Figure 1 5. This portion of the cardiac cycle is called diastole. The outlet valve is closed during diastole because arterial pressure is greater than intraventricular pressure. After the period of diastolic filling, the systolic phase of a new cardiac cycle is initiated. Excitation Efficient pumping action of the heart requires a precise coordination of the contraction of millions of individual cardiac muscle cells. Contraction of each cell is triggered when an electrical excitatory impulse (action potential) sweeps over its membrane. Proper coordination of the contractile activity of the individual cardiac muscle cells is achieved primarily by the conduction of action potentials from one cell to the next via gap junctions that connect all cells of the heart into a functional syncytium (ie, acting as one synchronous unit). In addition, muscle cells in certain areas of the heart are specifically adapted to control the frequency of cardiac excitation, the pathway of conduction, and the rate of the impulse propagation through various regions of the heart. The major components of this specialized excitation and conduction system are shown in Figure 1 6. They include the sino-atrial node (SA node), the atrioventricular node (AV node), the bundle of His, and the right and left bundle branches made up of specialized cells called Purkinje fibers. The SA node contains specialized cells that normally function as the heart's pacemaker and initiate the action potential that is conducted through the heart. The AV node contains slowly conducting cells that normally function to create a slight delay between atrial contraction and ventricular contraction. The Purkinje fibers are specialized for rapid conduction and assure that all ventricular cells contract at nearly the same instant. Requirements for Effective Operation For effective efficient ventricular pumping action, the heart must be functioning properly in five basic respects: - 1. The contractions of individual cardiac muscle cells must occur at regular intervals and be synchronized (not arrhythmic).
- 2. The valves must open fully (not stenotic).
- 3. The valves must not leak (not insufficient or regurgitant).
- 4. The muscle contractions must be forceful (not failing).
- 5. The ventricles must fill adequately during diastole.
In the subsequent chapters, we will study in detail how these requirements are met in the normal heart. Control of the Heart & Cardiac Output Diastolic Filling  One of the most fundamental causes of variations in stroke volume was described by William Howell in 1884 and by Otto Frank in 1894 and formally stated by E. H. Starling in 1918. These investigators demonstrated that as cardiac filling increases during diastole, the volume ejected during systole also increases. As a consequence, and as illustrated in Figure 1 7, with other factors equal, stroke volume increases as cardiac end-diastolic volume increases. This phenomenon (commonly referred to as Starling's law of the heart) is an intrinsic property of the cardiac muscle and is one of the primary regulators of cardiac output. The mechanisms responsible for this phenomenon will be described in detail in subsequent chapters. One of the most fundamental causes of variations in stroke volume was described by William Howell in 1884 and by Otto Frank in 1894 and formally stated by E. H. Starling in 1918. These investigators demonstrated that as cardiac filling increases during diastole, the volume ejected during systole also increases. As a consequence, and as illustrated in Figure 1 7, with other factors equal, stroke volume increases as cardiac end-diastolic volume increases. This phenomenon (commonly referred to as Starling's law of the heart) is an intrinsic property of the cardiac muscle and is one of the primary regulators of cardiac output. The mechanisms responsible for this phenomenon will be described in detail in subsequent chapters.
Autonomic Neural Influences While the heart can inherently beat on its own, cardiac function can be influenced profoundly by neural inputs from both the sympathetic and parasympathetic divisions of the autonomic nervous system. These inputs allow us to modify cardiac pumping as is appropriate to meet changing homeostatic needs of the body. All portions of the heart are richly innervated by adrenergic sympathetic fibers. When active, these sympathetic nerves release norepinephrine (noradrenaline) on cardiac cells. Norepinephrine interacts with 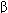 1-adrenergic receptors on cardiac muscle cells to increase heart rate, increase action potential conduction velocity, and increase force of contraction and rates of contraction and relaxation. Overall, sympathetic activation acts to increase cardiac pumping. 1-adrenergic receptors on cardiac muscle cells to increase heart rate, increase action potential conduction velocity, and increase force of contraction and rates of contraction and relaxation. Overall, sympathetic activation acts to increase cardiac pumping. Cholinergic parasympathetic nerve fibers travel to the heart via the vagus nerve and innervate the SA node, the AV node, and atrial muscle. When active, these parasympathetic nerves release acetylcholine on cardiac muscle cells. Acetylcholine interacts with muscarinic receptors on cardiac muscle cells to decrease heart rate (SA node) and decrease action potential conduction velocity (AV node). Parasympathetic nerves may also act to decrease the force of contraction of atrial (not ventricular) muscle cells. Overall, parasympathetic activation acts to decrease cardiac pumping. Usually an increase in parasympathetic nerve activity is accompanied by a decrease in sympathetic nerve activity, and vice versa. | | The Vasculature Blood that is ejected into the aorta by the left heart passes consecutively through many different types of vessels before it returns to the right heart. As diagrammed in Figure 1 8, the major vessel classifications are arteries, arterioles, capillaries, venules, and veins. These consecutive vascular segments are distinguished from one another by differences in physical dimensions, morphological characteristics, and function. One thing that all vessels have in common is that they are lined with a contiguous single layer of endothelial cells. In fact, this is true for the entire circulatory system including the heart chambers and even the valve leaflets. Some representative physical characteristics are shown in Figure 1 8 for each of the major vessel types. It should be realized, however, that the vascular bed is a continuum and that the transition from one type of vascular segment to another does not occur abruptly. The total cross-sectional area through which blood flows at any particular level in the vascular system is equal to the sum of the cross-sectional areas of all the individual vessels arranged in parallel at that level. The number and total cross-sectional area values presented in Figure 1 8 are estimates for the entire systemic circulation. Arteries are thick-walled vessels that contain, in addition to some smooth muscle, a large component of elastin and collagen fibers. Primarily because of the elastin fibers, which can stretch to twice their unloaded length, arteries can expand to accept and temporarily store some of the blood ejected by the heart during systole and then, by passive recoil, supply this blood to the organs downstream during diastole. The aorta is the largest artery and has an inside diameter of about 25 mm. Arterial diameter decreases with each consecutive branching, and the smallest arteries have diameters of approximately 0.1 mm. The consecutive arterial branching pattern causes an exponential increase in arterial numbers. Thus, while individual vessels get progressively smaller, the total cross-sectional area available for blood flow within the arterial system increases to severalfold that in the aorta. Arteries are often referred to as conduit vessels because they have relatively low and unchanging resistance to flow.  Arterioles are smaller and structured differently than arteries. In proportion to lumen size, arterioles have much thicker walls with more smooth muscle and less elastic material than arteries. Because arterioles are so muscular, their diameters can be actively changed to regulate the blood flow through peripheral organs. Despite their minute size, arterioles are so numerous that in parallel their collective cross-sectional area is much larger than that at any level in arteries. Arterioles are often referred to as resistance vessels because of their high and changeable resistance, which regulates peripheral blood flow through individual organs. Arterioles are smaller and structured differently than arteries. In proportion to lumen size, arterioles have much thicker walls with more smooth muscle and less elastic material than arteries. Because arterioles are so muscular, their diameters can be actively changed to regulate the blood flow through peripheral organs. Despite their minute size, arterioles are so numerous that in parallel their collective cross-sectional area is much larger than that at any level in arteries. Arterioles are often referred to as resistance vessels because of their high and changeable resistance, which regulates peripheral blood flow through individual organs.
Capillaries are the smallest vessels in the vasculature. In fact, red blood cells with diameters of 7 m must deform to pass through them. The capillary wall consists of a single layer of endothelial cells, which separate the blood from the interstitial fluid by only about 1 m. Capillaries contain no smooth muscle and thus lack the ability to change their diameters actively. They are so numerous that the total collective cross-sectional area of all the capillaries in systemic organs is more than 1000 times that of the root of the aorta. Given that capillaries are about 0.5 mm in length, the total surface area available for exchange of material between blood and interstitial fluid can be calculated; it exceeds 100 m2. For obvious reasons, capillaries are viewed as the exchange vessels of the cardiovascular system. In addition to the transcapillary diffusion of solutes that occurs across these vessel walls, there can sometimes be net movements of fluid (volume) into and/or out of capillaries. For example, tissue swelling (edema) is a result of net fluid movement from plasma into the interstitial space. After leaving capillaries, blood is collected in venules and veins and returned to the heart. Venous vessels have very thin walls in proportion to their diameters. Their walls contain smooth muscle and the diameters of venous vessels can actively change. Because of their thin walls, venous vessels are quite distensible. Therefore, their diameters change passively in response to small changes in transmural distending pressure (ie, the difference between the internal and external pressures across the vessel wall). Venous vessels, especially the larger ones, also have one-way valves that prevent reverse flow. As will be discussed later, these valves are especially important in the cardiovascular system's operation during standing and during exercise. It turns out that peripheral venules and veins normally contain more than 50% of the total blood volume. Consequently, they are commonly thought of as the capacitance vessels. More importantly, changes in venous volume greatly influence cardiac filling and therefore cardiac pumping. Thus, peripheral veins actually play an extremely important role in controlling cardiac output. Control of Blood Vessels  Blood flow through individual vascular beds is profoundly influenced by changes in activity of sympathetic nerves innervating arterioles. These nerves release norepinephrine from their endings which interacts with Blood flow through individual vascular beds is profoundly influenced by changes in activity of sympathetic nerves innervating arterioles. These nerves release norepinephrine from their endings which interacts with  -adrenergic receptors on the smooth muscle cells to cause contraction and thus arteriolar constriction. The reduction in arteriolar diameter increases vascular resistance and decreases blood flow. These neural fibers provide the most important means of reflex control of vascular resistance and organ blood flow. -adrenergic receptors on the smooth muscle cells to cause contraction and thus arteriolar constriction. The reduction in arteriolar diameter increases vascular resistance and decreases blood flow. These neural fibers provide the most important means of reflex control of vascular resistance and organ blood flow.
Arteriolar smooth muscle is also very responsive to changes in the local chemical conditions within an organ that accompany changes in the metabolic rate of the organ. For reasons to be discussed later, increased tissue metabolic rate leads to arteriolar dilation and increased tissue blood flow. Venules and veins are also richly innervated by sympathetic nerves and constrict when these nerves are activated. The mechanism is the same as that involved with arterioles. Thus, increased sympathetic nerve activity is accompanied by decreased venous volume. The importance of this phenomenon is that venous constriction tends to increase cardiac filling and therefore cardiac output via Starling's law of the heart. There is no important neural or local metabolic control of either the arterial or capillary vessels. | | Blood  Blood is a complex fluid that serves as the medium for transporting substances between the tissues of the body and performs a host of other functions as well. Normally about 40% of the volume of whole blood is occupied by blood cells that are suspended in the watery fluid, plasma, which accounts for the rest of the volume. The fraction of blood volume occupied by cells is a clinically important parameter termed the hematocrit: Blood is a complex fluid that serves as the medium for transporting substances between the tissues of the body and performs a host of other functions as well. Normally about 40% of the volume of whole blood is occupied by blood cells that are suspended in the watery fluid, plasma, which accounts for the rest of the volume. The fraction of blood volume occupied by cells is a clinically important parameter termed the hematocrit:
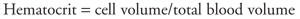
Blood Cells Blood contains three general types of "formed elements": red cells, white cells, and platelets (see Appendix A). All are formed in bone marrow from a common stem cell. Red cells are by far the most abundant. They are specialized to carry oxygen from the lungs to other tissues by binding oxygen to hemoglobin, an iron-containing heme protein concentrated within red blood cells. Because of the presence of hemoglobin, blood can transport 40 to 50 times the amount of oxygen that plasma alone could carry. In addition, the hydrogen ion buffering capacity of hemoglobin is vitally important to the blood's capacity to transport carbon dioxide. A small but important fraction of the cells in blood are white cells or leukocytes. Leukocytes are involved in immune processes. Appendix A gives more information on the types and function of leukocytes. Platelets are small cell fragments that are important in the blood-clotting process. Plasma Plasma is the liquid component of blood and, as indicated in Appendix B, is a complex solution of electrolytes and proteins. Serum is the fluid obtained from a blood sample after it has been allowed to clot. For all practical purposes, the composition of serum is identical to that of plasma except that it contains none of the clotting proteins. Inorganic electrolytes (inorganic ions such as sodium, potassium, chloride, and bicarbonate) are the most concentrated solutes in plasma. Of these, sodium and chloride are by far the most abundant and, therefore, are primarily responsible for plasma's normal osmolarity of about 300 mOsm/L. To a first approximation, the "stock" of the plasma soup is a 150 mM solution of sodium chloride. Such a solution is called isotonic saline and has many clinical uses as a fluid that is compatible with cells. Plasma normally contains many different proteins. Most plasma proteins can be classified as albumins, globulins, or fibrinogen on the basis of different physical and chemical characteristics used to separate them. More than 100 distinct plasma proteins have been identified and each presumably serves some specific function. Many plasma proteins are involved in blood clotting or immune/defense reactions. Many others are important carrier proteins for a variety of substances including fatty acids, iron, copper, vitamin D, and certain hormones. Proteins do not readily cross capillary walls and, in general, their plasma concentrations are much higher than their concentrations in the interstitial fluid. As will be discussed, plasma proteins play an important osmotic role in transcapillary fluid movement and thus the distribution of extracellular volume between the plasma and interstitial compartments. Albumin plays an especially strong role in this regard simply because it is by far the most abundant of the plasma proteins. Plasma also serves as the vehicle for transporting nutrients and waste products. Thus, a plasma sample contains many small organic molecules such as glucose, amino acids, urea, creatinine, and uric acid whose measured values are useful in clinical diagnosis. | | Foundation for Subsequent Chapters This first chapter has presented an overall description of the design of the cardiovascular system. Some important, basic, bottom-line principles that should help you understand many aspects of cardiovascular function have been included. (See the study questions at the end of this chapter, for example.) Subsequent chapters will expand these concepts in much greater detail, but we urge students not to lose sight of the overall picture presented in this chapter. It may be useful to repeatedly refer back to this material. | | Study Questions - 1 1. Which organ in the body always receives the most blood flow?
- 1 2. Whenever skeletal muscle blood flow increases, blood flow to other organs must decrease. True or false?
- 1 3. When a heart valve does not close properly, a sound called a murmur can often be detected as the valve leaks. Would you expect a leaky aortic valve to cause a systolic or diastolic murmur?
- 1 4. Slowing of action potential conduction through the AV node will slow heart rate. True or false?
- 1 5. Calculate cardiac output from the following data:
- Pulmonary arterial pressure = 20 mmHg
- Pulmonary venous pressure = 0 mmHg
- Pulmonary vascular resistance = 4 mmHg x min/L
- 1 6. a. Determine the vascular resistance of a resting skeletal muscle from the following data:
- Mean arterial pressure = 100 mmHg
- Mean venous pressure = 0 mmHg
- Blood flow to the muscle = 5 mL/min
- b. Assume that when the muscle is exercising, the resistance vessels dilate so that their internal radius doubles. If blood pressure does not change, what is the blood flow through the exercising muscle?
- c. What is the vascular resistance of this exercising skeletal muscle?
- 1 7. Usually an individual who has lost a significant amount of blood is weak and does not reason very clearly. Why would blood loss have these effects?
- 1 8. What direct cardiovascular consequences would you expect from an intravenous injection of norepinephrine?
- 1 9. What direct cardiovascular effects would you expect from an intravenous injection of a drug that stimulates -adrenergic receptors but not -adrenergic receptors?
- 1 10. Individuals with high arterial blood pressure (hypertension) are often treated with drugs that block -adrenergic receptors. What is the rationale for such treatment?
- 1 11. The clinical laboratory reports a serum sodium ion value of 140 mEq/L in a blood sample you have taken from a patient. What does this tell you about the sodium ion concentration in plasma, in interstitial fluid, and in intracellular fluid?
- 1 12. An individual has had the "flu" for 3 days with severe vomiting and diarrhea. How is this likely to influence his or her hematocrit?
See answers. | | Suggested Readings Handbook of Physiology Series. Section 2: The Cardiovascular System, New York, NY: Oxford University Press.
| Volume I: The Heart. Edited by Page E, Fozzard H, Solaro RJ. 2000.
| Volume II: Vascular Smooth Muscle. Edited by Bohr DF, Somlyo AP, Sparks HV, 1980.
| Volume III: Peripheral Circulation and Organ Blood Flow. Edited by Shepherd JT, 1983.
| Volume IV: Microcirculation. Edited by Renkin E, Michel CC, 1984.
| Harvey W. (1628). The Movement of the Heart and Blood. (translated by Witteridge G.) Oxford: Blackwell Scientific Publications; 1976.
| Rowell LB. Human Cardiovascular Control. New York, NY: Oxford University Press; 1993.
| | | | |