20 - Normal Heart
Editors: Mills, Stacey E.
Title: Histology for Pathologists, 3rd Edition
Copyright 2007 Lippincott Williams & Wilkins
> Table of Contents > VII - Alimentary Tract > 26 - Vermiform Appendix
26
Vermiform Appendix
Terry L. Gramlich
Robert E. Petras
Gross Anatomy/Surgical Perspective
The vermiform (wormlike) appendix is a slender tubular extension of the posteromedial aspect of the cecum originating below, and within 1 to 3 cm of, the ileocecal junction. Although the appendix has a relatively constant relationship with the cecum at the appendiceal base, the remainder of its length can be found in a variable number of positions, including retrocecal, subcecal, pelvic, and juxtaileal (1,2,3). A retrocecal position occurs most commonly, being present in nearly 70% of the population (3,4). Unusual locations, including a vermiform appendix buried within the cecal wall, have been documented (5). Although the appendix itself lacks taeniae, the base of the vermiform appendix lies at the convergence of the three cecal/ascending colon taeniae. These aid in locating the appendix when it is not readily apparent; the prominent anterior taenia is most easily traced for this purpose (1,6).
Vermiform appendices can vary remarkably in length but average 7 to 10 cm (2,4). The peritoneum covers almost all its external surface. The mesoappendix (mesentery of the appendix), a fold of peritoneum contiguous with the mesentery of the terminal ileum, extends along its length, terminating just proximal to the tip (1).
The appendiceal vascular supply courses within the mesoappendix; and, with distal progression, these vessels gradually rest nearer to the appendiceal muscular wall. In the proximity of the tip where there is no mesoappendix, blood vessels lie essentially unprotected on its external surface (1). The appendicular artery, a derivative of the inferior branch of the ileocolic artery of the superior mesenteric trunk, provides the majority of blood to the appendix (4,7). However, a variable supply with accessory arterial contributions is not unusual (8). Branches of the ileocolic vein drain the appendiceal venous network into the superior mesenteric vein and eventually into the portal circulation, whereas lymphatic vessels drain into regional (e.g., ileocolic) lymph nodes (6). Innervation is derived from branches of the vagus
P.650
nerve (parasympathetic) and superior mesenteric plexus (sympathetic). Venous, lymphatic, and neural components closely follow the arterial vasculature (6).
Grossly, the external surface of the vermiform appendix appears smooth, pink-tan or gray, and glistening. The appendiceal diameter typically measures 5 to 8 mm. The wall is tan-white and the mucosal lining is light yellow, often disclosing a nodular appearance imparted by the characteristic and prominent lymphoid component (9). Because of these lymphoid aggregates, the central lumen on cross section is often irregular (stellate) rather than round. The normal luminal diameter measures 1 to 3 mm; however, in one study a luminal diameter of 1.2 cm or more was arbitrarily defined as dilatation (10). Focal occlusions of the appendiceal lumen are not uncommon (9).
Development of the Vermiform Appendix and Congenital Anomalies
The vermiform appendix originates from the primordial structure termed the cecal diverticulum (5,11). First apparent during the sixth week of fetal life, this blind-ended sac progressively develops. Its most proximal portion, in continuity with the remainder of the large bowel, enlarges and expands, forming the cecum proper, whereas its distal aspect or apex simply elongates, remains narrow, and becomes the vermiform appendix (11). Continued growth through infancy and childhood leads to differing cecoappendiceal relationships over this period. For example, the infantile cecoappendiceal junction lacks a conspicuous transition; the appendix arises from the inferior aspect of the cecum in this age group. In contrast, an abrupt, easily recognizable junction on the posteromedial cecum is observed in the adult (2).
Abnormal embryologic development can result in agenesis, hypoplasia, and various duplications or even triplication of the appendix (5,9,12,13,14). Duplication of the appendix can mimic cecal duplication. In general, appendiceal duplication is recognized by the presence of complete and separate inner circular and outer longitudinal muscle bands and the presence of a prominent lymphoid component (12).
Duplications have been well described and categorized and can be associated with other complex and life-threatening congenital anomalies. The classification of appendiceal duplications includes type A, an appendix with a common base, single cecum, and bifurcated distal portion; type B, two separate appendices with distinct bases arising from a single cecum; and type C, two cecal structures, each with its own single appendix (12,13). The type C anomaly is always associated with other organ duplications and often necessitates extensive operative correction in infancy; a type B variant is also associated with other systemic anomalies (12). However, the majority of type B and all type A duplications are found incidentally or during operation for suspected appendicitis in older children and adults.
Function
The exact role of the appendix is uncertain. However, rather than simply representing a vestigial, functionless structure, the abundant quantity of organized lymphoid tissue suggests involvement in mucosal immunity (15). It has been suggested that B lymphocytes derived from the appendix migrate and populate distant sites of the gastrointestinal tract lamina propria and evolve in these widespread foci into functional immunoglobulin (Ig)A-secreting plasma cells (15,16). In this role, the appendix can both attenuate potentially harmful immunoglobulin responses and enhance regional mucosal immunity (16).
Normal Histology of the Appendix
The histologic composition of the appendix is similar to that of the large bowel. The four layers, from its luminal to external surface, include the mucosa, submucosa, muscularis externa (or propria), and serosa. The distinctive features of the appendix are emphasized.
Mucosal Architecture and Design
A single layer of surface epithelium covers the luminal aspect of the appendiceal mucosa. This overlies the lamina propria within which crypts, or intestinal glands, contiguous with the surface epithelial cells are irregularly dispersed (Figure 26.1). The lamina propria is a cellular layer with an abundant migratory cell component and prominent, often
P.651
confluent, lymphoid aggregates. In contrast to the scattered lymphoid nodules within the large bowel proper, the appendix, particularly in young individuals, contains abundant and organized lymphoid structures spread around its entire luminal circumference. These lymphoid nodules often distort the luminal contour (9,17). The outermost component and limit of the mucosa is the muscularis mucosae. This slender fibromuscular band is poorly developed in the appendix and often focally deficient.
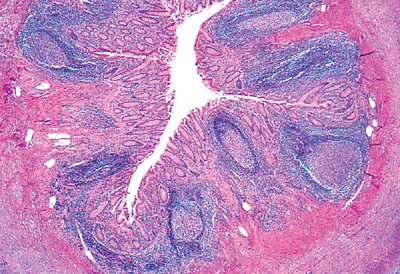 |
Figure 26.1 Low-magnification view of a cross section of the vermiform appendix. The irregular (stellate) lumen is lined by a single layer of surface epithelium. The remainder of the mucosa (crypts, surrounding lamina propria, and the rather inconspicuous muscularis mucosae) surrounds this surface epithelial layer. Note the characteristic lymphoid nodules within the lamina propria that also extend into the submucosa. |
Surface Epithelium
Several different cell types comprise the surface epithelium. A prominent cell that can be identified at the light microscopic level is tall and columnar with eosinophilic cytoplasm, and it has a round, basally located nucleus (Figure 26.2). These cells represent several distinct cell types that can be differentiated at the ultrastructural level, including senescent mucous cells, so-called absorptive cells, and membranous or M cells (18,19,20,21). Goblet cells with distinctive apical mucin droplets surrounded by eosinophilic cytoplasm and undermined by an attenuated basal nucleus intermix with the columnar cells (Figure 26.2). The goblet cell apical mucin droplet contains both periodic acid-Schiff (PAS)-positive neutral mucin and Alcian blue positive acid sulfomucin. This combination results in the formation of a blue-purple color in a mixed Alcian blue/PAS stain (19,22) (Figure 26.3). Overlying lymphoid aggregates, as in other portions of the small and large bowel, is a specialized or follicle-associated epithelium that is distinct from the surrounding surface epithelium. It characteristically has fewer goblet cells, and many of the columnar cells are of the M-cell type (22) (Figure 26.4). The M cell, a specialized epithelial cell, assists in luminal transport of antigens into the epithelium for appropriate immunologic processing (23,24); M cells are columnar in shape with an attenuated brush border; several lymphocytes are often seen deforming their dependent cytoplasm. Definitive characterization rests on ultrastructural examination, which shows apical cytoplasmic vesicles and shortened microvilli or microfolds (21,23). Because circumferentially distributed organized lymphoid aggregates and lymphoid tissue are prominent in the normal appendix, the specialized follicle-associated epithelium often lines the majority of the appendiceal lumen. Thus, functionally, the surface epithelium is probably primarily involved in antigen processing, as well as in forming a barrier to luminal contents. The luminal surface is also the site where senescent cells are sloughed into the lumen (19,20). Scattered endocrine cells can be seen within the surface epithelium but are more abundant in the underlying crypts. Migratory T and B lymphocytes can be found anywhere within the surface epithelium (25,26) but are more abundant in the follicle-associated epithelium (Figure 26.4).
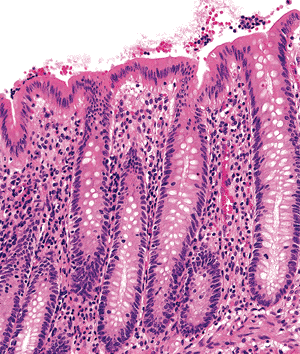 |
Figure 26.2 The surface epithelium is composed of a single layer of predominantly columnar cells with rare interspersed goblet cells. The crypt linings have a similar cellular composition but contain more goblet cells. |
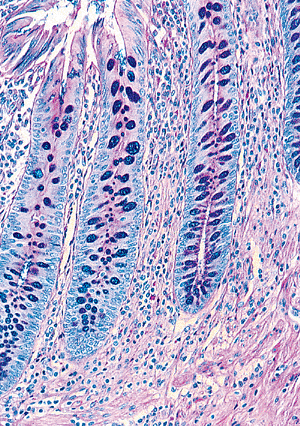 |
Figure 26.3 Because goblet cells contain both neutral and acid mucopolysaccharides, their apical mucin droplets stain blue-purple with the mixed Alcian blue/PAS preparation. |
P.652
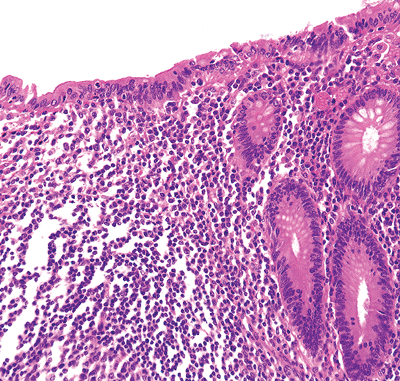 |
Figure 26.4 Surface epithelium overlying a lymphoid aggregate composed solely of tall columnar cells without intermixed goblet cells. Ultrastructurally, most of these would be classified as membranous or M cells. Note the increased numbers of intraepithelial lymphocytes between the individual columnar cells. Directly beneath the epithelium is the dome region of the lymphoid nodule. The apical portion of the germinal center with surrounding mantle zone is present near the bottom of the micrograph. |
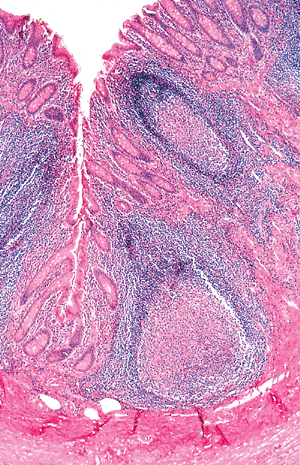 |
Figure 26.5 Lymphoid aggregates are often a prominent component within the appendiceal mucosa. Note the absence of crypts in the region of the lymphoid nodules and the distortion of surrounding crypts. This is a normal finding in the appendix and is similar to the alteration associated with isolated lymphoid aggregates in the colon. |
Crypt Epithelium
In contrast to the colon, where crypts line up evenly like test tubes in a rack, appendiceal crypts are more irregular in shape, length, and distribution (27). In areas with abundant lymphoid tissue or lymphoid aggregates, crypts are typically absent (28) (Figure 26.5).
Several different cell types line the crypts. The goblet and columnar cell variants discussed above are the most abundant (Figures 26.2, 26.6). Undifferentiated stem cells are scattered about but are inconspicuous. These are typically located at the crypt base, rest on the basement membrane, and do not extend to the crypt lumen; they are best identified by ultrastructural means (19). Isolated or clustered endocrine cells are seen along the crypt epithelium. Their appearance varies from a flask-shaped cell with a narrow strip of apical cytoplasm contiguous with the surface to a spindle-shaped cell with no luminal connection (29,30). Although some endocrine cells can be recognized on hematoxylin and eosin (H&E) stained sections by their eosinophilic, infranuclear granules (31) (Figure 26.6),
P.653
definitive identification rests on immunohistologic analysis for chromogranin (or other pan-reactive neuroendocrine marker) (Figure 26.7) or ultrastructural analysis, which discloses neurosecretory granules within their cytoplasm. More specific immunohistologic methods show that endocrine cells within the appendiceal epithelium contain a variety of amine and polypeptide substances, including serotonin, substance P, somatostatin, and enteroglucagon (32). Paneth's cells also can be found in the crypt bases within the normal appendix in nearly 96% of specimens (33,34,35). This cell has a basally situated, round nucleus with a conspicuous nucleolus and abundant eosinophilic supranuclear granules (Figure 26.8); their function remains unknown, but they probably play a role in microbial regulation (33).
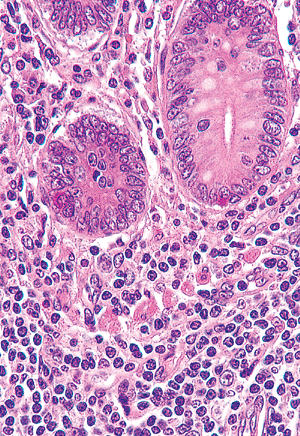 |
Figure 26.6 Crypts lying within a normocellular lamina propria. The round or ovoid crypts are lined predominantly by eosinophilic columnar cells and goblet cells. A single endocrine cell (containing infranuclear eosinophilic granules) is present at the base of each crypt. The lamina propria contains plasma cells, lymphocytes, and scattered eosinophils. Note the polygonal cells with abundant eosinophilic cytoplasm within the lamina propria. These are the subepithelial (laminal propria) endocrine cells that are often found near the crypt bases. |
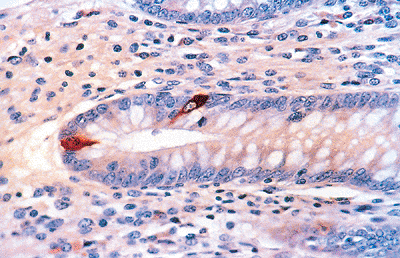 |
Figure 26.7 Scattered endocrine cells within the epithelium of an appendiceal crypt highlighted with antichromogranin. Intense red-brown cytoplasmic staining is evident in these endocrine cells. |
Intraepithelial lymphocytes occur within the crypt epithelium (36,37), but neutrophils and plasma cells are not normal constituents of either epithelial compartment. Rarely, gastric, ileal, or esophageal squamous-type mucosa can be seen interrupting the normal appendiceal lining; some recognize these as true heterotopias (38,39,40).
The crypt functions in cell production and renewal because all cells of both epithelial compartments originate from the crypt's stem cells. Most of these cells travel to the surface epithelium, where they are subsequently sloughed intraluminally; the exception (Paneth's cell) remains in the crypt base (36,37). It is believed that apoptosis within the crypt probably functions to regulate cell migration toward the surface; however, it is uncertain whether this type of cell death is responsible for epithelial cell loss into the lumen (41).
Subepithelial Basement Membrane
A slender zone separates the epithelial compartments from the lamina propria and is composed of collagen and other matrix components (42). The subepithelial basement membrane stabilizes the epithelial layers. A PAS stain can be used to highlight this layer, which measures only microns in thickness (20,42) (Figure 26.3).
Lamina Propria
The lamina propria, the central layer of the mucosa, surrounds the crypts and forms a connective tissue framework around them. Its structural components are collagen and elastic fibers and associated fibroblasts intermingled with blood capillaries, lymphatics, and nerve fibers (18,19,20). As in the large bowel, its migratory cell component consists primarily of plasma cells and T lymphocytes, along with scattered macrophages, eosinophils, B lymphocytes, and mast cells (18,26,43) (Figures 26.6, 26.8). However, depending on an individual's age, a varying number of organized lymphoid nodules distort the lamina proprial architecture. These lymphoid aggregates can extend beneath the muscularis mucosae into the underlying submucosa (Figures 26.1, 26.5), are often confluent, and appear similar in composition and function to the Peyer's patches of the small bowel (21). As in Peyer's patches (see Chapter 24), this lymphoid network of the appendix is compartmentalized into (a) follicle, (b) dome, (c) interfollicular (or parafollicular) region, and (d) follicle-associated epithelium (44,45,46) (Figure 26.9). The follicle has, in most cases, a germinal center containing a polymorphic cellular population of small and large B lymphocytes in various stages of maturation, occasional CD4+ T-helper cells, and tingible body macrophages; these reactive centers invariably contain mitoses (15,45,46,47) (Figure 26.10). Immediately surrounding the germinal center is the mantle zone, a darkly staining cuff of small, round B lymphocytes. Overlying the
P.654
lymphoid aggregate and beneath the epithelium is the dome region (mixed cell zone), which is composed of a heterogeneous population of cells including B and T lymphocytes, macrophages, and occasional plasma cells (21,46). Both the lymphoid aggregate and the dome region are supported by a structural framework provided by dendritic reticulum cells and their processes (16,47). A prominent collagenous network and closely associated lymphatic vessels surround and define the lymphoid nodule (16). This collagenous/fibrous border is contiguous with the connective tissue framework of the interfollicular zones and adjacent lamina propria (16). The zone surrounding a single lymphoid nodule (parafollicular region) and the area between confluent lymphoid aggregates (the interfollicular region) consist predominantly of T lymphocytes (46) (Figure 26.11). Moreover, the ratio of T-helper/inducer (CD4+) to T-suppressor/cytotoxic (CD8+) lymphocytes is normally about 8:1 in these T-lymphocyte rich areas (46). Finally, as detailed previously, the overlying epithelium is specialized and distinct from the usual surface epithelium.
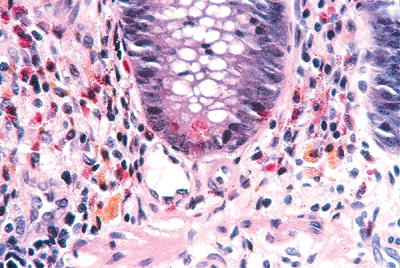 |
Figure 26.8 Appendiceal crypt disclosing Paneth's cell (at its base) with characteristic supranuclear eosinophilic granules. The surrounding lamina propria has a conspicuous, albeit normal, quantity of eosinophils. Also, note the golden brown, granular pigment within the macrophages, which is characteristic of melanosis. |
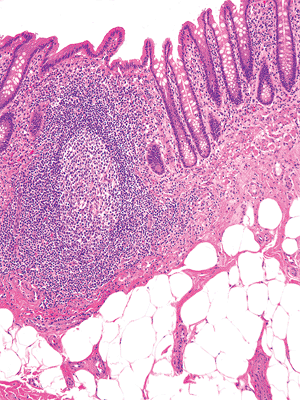 |
Figure 26.9 Characteristic lymphoid nodule within lamina propria of appendix. A germinal center forms the core of the follicle and is surrounded, at least in part, by a mantle zone of small round lymphocytes. Between the overlying epithelium and the mantle is the dome, which contains a mixed cellular population of lymphocytes, plasma cells, and macrophages. A portion of the parafollicular area (T-lymphocyte zone) is seen. Lymphatic and blood vessels are seen beneath the lymphoid nodule in the underlying superficial submucosa. |
The immunophenotypic cellular composition of the appendiceal mucosa is different from the colon. Although the quantity of lymphoid and plasma cells containing IgA and IgM is similar in both, IgG-containing cells are more abundant in the appendix (15,46) (Figure 26.12). In fact, nearly 50% of the those along the follicle borders, including the dome region, are IgG immunoreactive, whereas IgA-containing cells are more abundant in distant lamina proprial sites (15).
Lymphoid tissue, although a characteristic feature of the appendix, varies in quantity with age. The newborn's appendix contains scant or no lymphoid tissue. With increasing age the lymphoid nodules accumulate, peaking in the first decade (17,48). Lymphoid aggregates then steadily diminish in quantity throughout the remainder of life. However, appendices excised incidentally from middle-aged adults can still occasionally show a prominent organized lymphoid component (10). In contrast, lymphoid nodules and associated lymphocytes can be scant in the central obliterative form of appendiceal neuroma (fibrous obliteration of the appendiceal lumen) and occasionally in appendices removed from normal patients at any age (10). Thus, a great range of normal variation exists in the appendix with respect to its lymphoid content.
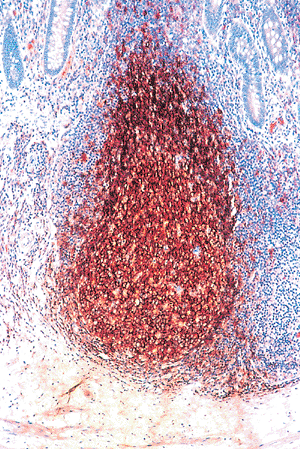 |
Figure 26.10 The germinal center and mantle zone contain predominantly B-lymphocytes. A pan B-lymphocyte immunomarker, L26, discloses this characteristic immunophenotype. Scattered macrophages and occasional T lymphocytes (see Figure 26.11) are also normally found within the germinal center. Only scattered B-lymphocytes are present within the interfollicular zone and adjacent lamina propria. |
P.655
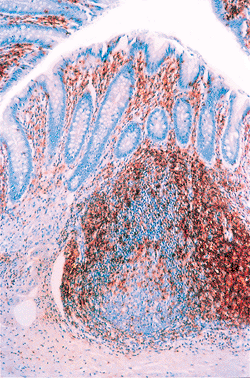 |
Figure 26.11 A pan T-cell immunomarker, Leu-22 (CD43), disclosing the characteristic T-lymphocyte distribution within the appendiceal mucosa. The lamina propria and interfollicular regions (between lymphoid follicles) are normally populated by numerous T lymphocytes. There is a sprinkling of T lymphocytes within the germinal center; these are predominantly T-helper/inducer (CD4+) lymphocytes. |
Histiocytes with intracellular golden-brown pigment (lipofuscin), not infrequently observed in the colonic mucosa, can also be found in the appendiceal lamina propria; this alteration results from anthracene-containing laxative abuse and has been termed melanosis coli when seen in the colon proper (9,49) (Figure 26.8). Interestingly, this pigmentation is a result of apoptosis induced by anthraquinones (41).
The lamina propria of the appendix contains a well-developed mucosal nervous plexus that is different from the more prominent submucosal and myenteric plexuses. Although all contain neurons (ganglion cells), Schwann cells, and neural processes (axons and neuropil), only the mucosal plexus contains endocrine (neurosecretory) cells. As a consequence, this network has been termed the mucosal neuroendocrine complex (50). These complexes, located just beneath the crypts, are composed of collections of endocrine cells and seen on H&E stained preparations as polygonal cells with pale granular cytoplasm (Figure 26.6), often intimately associated with spindled Schwann cells, neural processes, and occasional neurons. These collections, or neuroendocrine ganglia, are interconnected by neural fibers that can be highlighted immunohistologically with antibody preparations to neuron-specific enolase and, in a subset, to substance P (32); anti S-100 also can outline this network as it marks the accompanying Schwann cells. The mucosal plexus also communicates with other neural networks of the enteric nervous system (50,51,52,53). The subepithelial endocrine cells are not always conspicuous but can be highlighted using general neuroendocrine immunomarkers, such as antichromogranin (Figure 26.13) and antineuron-specific enolase, or by using electron microscopy (54,55). Most of these cells have been shown by specific immunohistologic analysis to contain serotonin (51,55). The mucosal neuroendocrine complex is believed to modulate neural communication, through serotonin mediators, between the epithelium and the deeper submucosal and intermuscular plexuses (55). Interestingly, because most appendiceal carcinoids are biphasic, consisting of an admixture of endocrine cells and S-100+ Schwann cells (similar to the architecture of the mucosal neuroendocrine complex), the majority of these appendiceal neoplasms are believed to be derived from these lamina propria endocrine cells rather than from the epithelium-based ones (51).
Muscularis Mucosae
The muscularis mucosae is a thin band of fibromuscular tissue separating the lamina propria and mucosal epithelium from the underlying submucosa. It characteristically forms a continuous layer in the large bowel (18); but, in the appendix, the muscularis mucosae is attenuated, poorly developed, and often focally absent, particularly in the region of penetrating lymphoid aggregates (28,56) (Figure 26.14). In these areas the muscularis mucosae may exist solely as isolated smooth muscle cells in the underlying submucosa (56).
Submucosa
The submucosa separates the mucosa from the muscularis externa. Its loose architectural framework contains a meshwork of collagenous and elastic fibers and associated fibroblasts (Figure 26.15). The submucosa can also contain inconspicuous migratory cells, such as macrophages, lymphoid and plasma cells, and mast cells, along with adipose tissue (17,57) (Figure 26.14). The morphologic appearance of the appendiceal submucosa and its primary role in maintaining structure are similar throughout the gastrointestinal tract (57). Arterioles, venules, blood capillaries, and lymphatic vessels are a prominent component of the submucosa (7,18) (Figure 26.15). Lymphatic vessels (or sinuses) are most prominent just beneath the bases of lymphoid aggregates (16). Neural structures, particularly Meissner's
P.656
plexus, are also conspicuous (Figure 26.16). This plexus consists of ganglia, collections of neurons (ganglion cells) with associated neuronal processes, and Schwann cells that interconnect, creating a neural network throughout the submucosal layer (58,59). The ganglion cell is large and oval with abundant eosinophilic cytoplasm; its vesicular nucleus is often eccentrically placed and contains a prominent nucleolus. The surrounding spindle and wavy Schwann cell component of the ganglia is less conspicuous on H&E stained preparations but can be highlighted with anti S-100 (Figure 26.16).
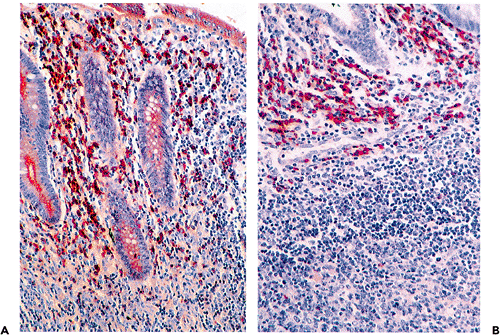 |
Figure 26.12 A. Immunohistologic preparation showing abundant IgA-containing plasma cells within lamina propria; the epithelial staining is a consequence of the secretory nature of the IgA molecule. B. Abundant IgG-bearing cells are characteristically located within the dome region and along the margins of lymphoid nodules in the appendix. |
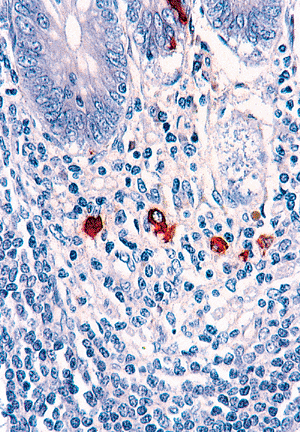 |
Figure 26.13 Antichromogranin highlights the subepithelial (lamina propria) endocrine cells beneath the crypts. These are more prominent and abundant in the appendix than in any other portion of gastrointestinal tract. Note also the epithelial-based endocrine cell in the overlying crypt. |
Muscularis Externa, Subserosal Region, and the Serosa
The thick smooth muscle layer lying between the submucosa and serosal portions of the appendix is the muscularis externa (or muscularis propria). It is separated into an inner circular layer and an outer longitudinal band (28). The individual smooth muscle cells are oval with blunted ends and form bundles of varying size. Occasionally, granular degeneration (eosinophilic cytoplasmic granularity) of individual or groups of smooth muscle cells is seen, particularly within the inner circular layer (56,60). Between the two muscle bands lies the myenteric (Auerbach's) plexus, which is similar morphologically and functionally to the previously described submucosal plexus of Meissner (59) (Figure 26.17).
P.657
Additionally, blood and lymphatic vessels and nerve fibers course through this muscular layer (16). Just external to the outer longitudinal smooth muscle layer is the subserosal region, consisting of loose connective tissue and ramifying blood vessels, lymphatics, and nerves. The exteriormost surface, or serosa, is lined by a single layer of cuboidal mesothelial cells that overlies a slender band of fibrous tissue. Only the attachment of the fibrofatty mesoappendix lacks a serosa (1).
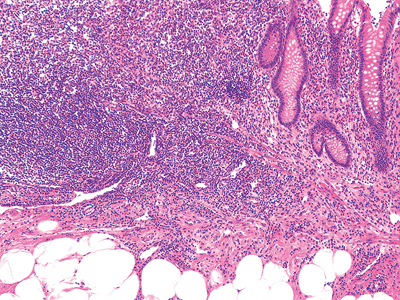 |
Figure 26.14 Characteristic focal deficiency of muscularis mucosae in region of lymphoid nodule. There is adipose tissue within the submucosa; this is a normal finding. |
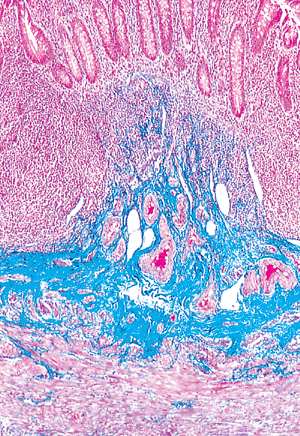 |
Figure 26.15 Normal appendiceal submucosa outlined in blue, highlighting its prominent collagenous framework. Numerous vascular spaces are also present within this layer. The mucosa (crypts) is above, and the inner circular layer of the muscularis externa is below (Masson's trichrome). |
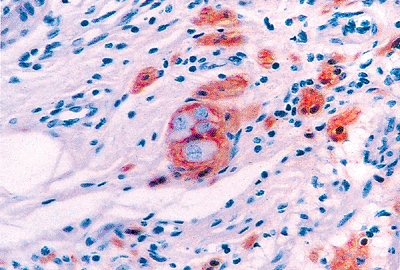 |
Figure 26.16 Submucosal neural network outlined with anti S-100. A single ganglion of Meissner's plexus is at the center; the ganglion cells (neurons) have abundant pale cytoplasm, a large eccentric nucleus, and show no immunoreactivity. The Schwann cells of the ganglion and those ensheathing the neuronal processes of the remainder of the plexus are highlighted. |
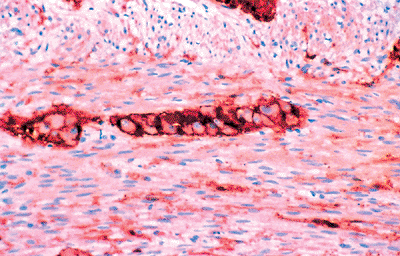 |
Figure 26.17 Anti S-100 highlighting Schwann cells of the neural network of the muscularis externa and a ganglion of the myenteric (Auerbach's) plexus. |
P.658
Special Considerations
Normal Variation of Mucosal Inflammation Versus Acute Appendicitis
Acute appendicitis is usually characterized by an abundant neutrophilic and eosinophilic infiltrate within the mucosa, submucosa, and often muscularis externa with at least focal mucosal ulceration; frequently suppurative inflammation extends into and through the appendiceal wall (9,10). However, the changes seen in early appendicitis can be quite minimal, and criteria considered sufficient to diagnose early acute appendicitis have varied (9,10,61,62,63,64,65,66). We agree that reactive lymphoid follicles are not a reliable sign of acute appendicitis (9). Focal collections of neutrophils within the lumen and lamina propria have been considered nondiagnostic by some investigators because many incidental appendectomy specimens contain these changes (9,10,61,64,65,66). However, we believe that if care is taken to recognize marginating neutrophils and early mucosal migration of these acute inflammatory cells (i.e., a result of the operative procedure alone), then other collections of neutrophils within the mucosa or intraluminal pus reflect stasis, infection, and changes of early appendicitis (62,63,64). Whether acute appendicitis becomes chronic or whether it can be recognized in a chronic state has long been debated (63). Fibrous obliteration of the appendiceal lumen is probably not a sequelae of acute appendicitis (53). However, prominent fibrosis, a marked chronic inflammatory cell infiltrate within the wall, and granulation tissue are abnormal and suggest an organizing appendicitis (9). Occasional specimens exhibit infiltration of the appendiceal wall by eosinophilic leukocytes with no other apparent abnormality (10). This change could reflect appendicitis elsewhere in the specimen that was not sampled; however, it remains possible that an infiltrate composed predominantly of eosinophils could represent appendicitis in a resolving phase or be a manifestation of eosinophilic gastroenteritis (62,67,68).
Obliteration of the Appendiceal Lumen (Appendiceal Neuromas)
Obliteration of the appendiceal lumen with absence of the lining mucosa and underlying crypts frequently occurs and has a prevalence in surgical specimens of nearly 30% (9,53). This process usually affects the distal aspect or just the tip, but occasionally the entire lumen is obliterated. This process is often termed fibrous obliteration; however, more recent studies have shown that in some cases the occlusive proliferation appears to be predominantly neurogenic (32,53,69). Other diagnostic terms have been proposed, including neurogenic appendicopathy and appendiceal neuroma. The typical appendiceal neuroma, or the central obliterative form, is composed of a collection of spindle cells in a loose myxoid background with varying amounts of collagen, fat, and chronic inflammatory cells (Figures 26.18,26.19). This typically occludes the lumen and blends imperceptibly with the surrounding submucosa (53). The involved segment usually lacks a mucosa, and lymphoid follicles are typically not seen (20). Immunostaining for neuron-specific enolase and S-100 highlights the spindle cells and identifies their neuronal (axons) and perineuronal (Schwann cell) nature, respectively (32,53) (Figure 26.20). Moreover, endocrine cells visualized with antineuron-specific enolase and antichromogranin (Figure 26.21) occur in many of the cases, usually intermingled with the other elements; serotonin and somatostatin have been identified in some of these endocrine cells by immunohistologic methods (32,53). Ultrastructural analysis discloses neuronal
P.659
processes, Schwann cells, and cells with neurosecretory granules (endocrine cells) corroborating the immunostaining results (53).
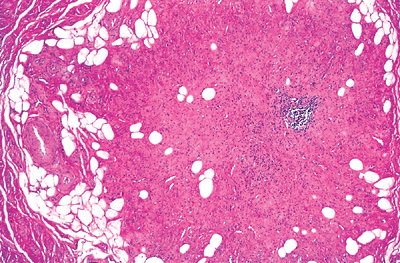 |
Figure 26.18 Obliteration of appendiceal lumen. The occlusive proliferation is composed of spindled cells within a collagenous and myxoid background, along with scattered adipocytes. A focus of chronic inflammatory cells is also present. |
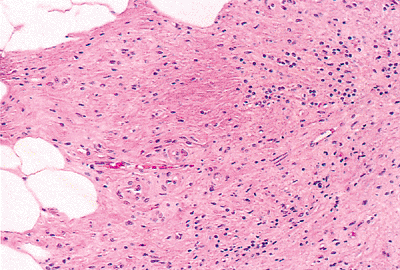 |
Figure 26.19 High magnification of Figure 26.18 showing spindled cell proliferation in an eosinophilic, fibromyxoid background. |
Another variant of this entity, the intramucosal appendiceal neuroma, primarily affects the mucosa, causing no luminal obliteration. Although morphologically similar to the central obliterative form, this intramucosal variant deceptively expands the lamina propria, separates the crypts, and replaces the usual prominent migratory cell population (53) (Figure 26.22). Immunostaining with S-100 can be helpful in visualizing these more subtle changes.
Both of these entities are believed to be proliferative rather than involutional, progressing through consecutive stages of growth, regression, and finally an end-stage with fibrosis (53,70). Overlapping features are therefore expected with varied admixtures of neurogenic components, collagen, and fat. It is hypothesized that associated endocrine cell hyperplasia, often found in adjacent uninvolved appendiceal segments, may be responsible for painful stimuli mimicking typical acute appendicitis (53). However, appendiceal neuromas are often found in specimens removed at incidental appendectomy.
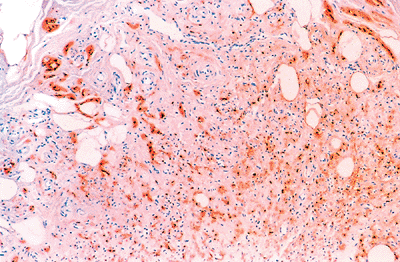 |
Figure 26.20 Prominent neurogenic (Schwann cell) component highlighted by anti S-100 within the obliterated lumen. |
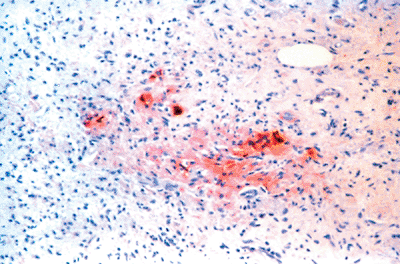 |
Figure 26.21 Scattered endocrine (neurosecretory) cells are evident within the obliterative luminal proliferation as highlighted by antichromogranin; specific immunomarkers show some of these to contain serotonin or somatostatin. |
Mucocele of the Appendix
The term mucocele has been used to describe a dilated appendiceal lumen filled with mucin (71). Mucocele, however, should not be used as a specific diagnostic term because the condition is almost always caused by a neoplastic proliferation, either a mucinous cystadenoma or mucinous cystadenocarcinoma (63,71,72). Characteristic architectural and cytologic features should permit identification of these entities.
 |
Figure 26.22 Intramucosal variant of appendiceal neuroma. The characteristic subtle spindle cell (schwannian) proliferation expands the lamina propria and separates the crypts. A diminished number of migratory cells is evident in this area. |
P.660
Dissection and Processing Techniques
Gross dissection and processing of the appendix are generally straightforward. Routine description of size, appearance, and any unusual lesions should be recorded. Luminal patency should be assessed (i.e., obliteration or dilatation) along with the focality and regional distribution of any changes. The tip should be closely inspected for carcinoid tumors because these commonly occur in the distal portion of the appendix (9,73). When grossly evident, they often appear as bulbous, tan-yellow expansions or nodules. However, a routine section of the tip is standard at most institutions and will identify small, grossly unidentifiable tumors (9). The common recommendation of a longitudinal section of the distal several centimeters is often difficult to orient, and we prefer a cross section of the tip. In the usual specimen, 1-cm serial cross-sectioning is performed along the entire length of the appendix. Two cross sections, one from the middle and one of the proximal line of resection, should be submitted for embedding. Because neoplastic proliferations of the appendix (e.g., mucinous cystadenoma/cystadenocarcinoma, carcinoid tumor, and its variants) are not infrequently discovered incidentally during microscopic evaluation of the specimen, we routinely sample the margin of resection. Otherwise, it could be difficult to reconstruct the gross specimen in an attempt to assess the adequacy of excision. The choice of a fixative is not crucial. However, we prefer the superior nuclear detail afforded by Hollande's solution over routine 4% formaldehyde solution. Modifications of dissection and processing may be necessary in certain situations.
References
1. Williams PL, Warwick R, Dyson M, Bannister LH, eds. Gray's Anatomy of the Human Body. 37th ed. New York: Churchill Livingstone; 1989.
2. Buschard K, Kjaeldgaard A. Investigation and analysis of the position, fixation, length, and embryology of the vermiform appendix. Acta Chir Scand 1973;139:293 298.
3. Wakeley CPG. The position of the vermiform appendix as ascertained by an analysis of 10,000 cases. J Anat 1933;67:277 283.
4. Thorek P. Anatomy and Surgery. 3rd ed. New York: Springer-Verlag; 1985.
5. Abramson DJ. Vermiform appendix located within the cecal wall. Anomalies and bizarre locations. Dis Colon Rectum 1983;26:386 389.
6. Hollinshead WH, Rosse C. Textbook of Anatomy. 4th ed. New York: Harper & Row; 1985.
7. Parks DA, Jacobson ED. Physiology of the splanchnic circulation. Arch Intern Med 1985;145:1278 1281.
8. Solanke TF. The blood supply of the vermiform appendix in Nigerians. J Anat 1968;102(pt 2):353 361.
9. Gray GF Jr, Wackym PA. Surgical pathology of the vermiform appendix. In: Sommers SC, Rosen PP, Fechner RE, eds. Pathology Annual. Part 2. Norwalk, CT: Appleton-Century-Croft; 1986:111 144.
10. Butler C. Surgical pathology of acute appendicitis. Hum Pathol 1981;12:870 878.
11. Moore KL. The Developing Human: Clinically Oriented Embryology. 3rd ed. Philadelphia: WB Saunders; 1982.
12. Bluett MK, Halter SA, Salhany KE, O'Leary JP. Duplication of the appendix mimicking adenocarcinoma of the colon. Arch Surg 1987;122:817 820.
13. Wallbridge PH. Double appendix. Br J Surg 1962;50:346 347.
14. Tinckler LF. Triple appendix vermiformis a unique case. Br J Surg 1968;55:79 81.
15. Bjerke K, Brandtzaeg P, Rognum TO. Distribution of immunoglobulin producing cells is different in normal human appendix and colon mucosa. Gut 1986;27:667 674.
16. Bockman DE. Functional histology of appendix. Arch Histol Jpn 1983;46:271 292.
17. Hwang JMS, Krumbhaar EB. The amount of lymphoid tissue of the human appendix and its weight at different age periods. Am J Med Sci 1940;199:75 83.
18. Hamilton SR. Structure of the colon. Scand J Gastroenterol Suppl 1984;93:13 23.
19. Shamsuddin AM, Phelps PC, Trump BF. Human large intestinal epithelium: light microscopy, histochemistry, and ultrastructure. Hum Pathol 1982;13:790 803.
20. Levine DS, Haggitt RC. Normal histology of the colon. Am J Surg Pathol 1989;13:966 984.
21. Bockman DE, Cooper MD. Early lymphoepithelial relationships in human appendix: a combined light- and electron-microscopic study. Gastroenterology 1975;68(pt 1):1160 1168.
22. Filipe MI. Mucins in the human gastrointestinal epithelium: a review. Invest Cell Pathol 1979;2:195 216.
23. Owen RL, Jones AL. Epithelial cell specialization within human Peyer's patches: an ultrastructural study of intestinal lymphoid follicles. Gastroenterology 1974;66:189 203.
24. Wolf JL, Bye WA. The membranous epithelial (M) cell and the mucosal immune system. Annu Rev Med 1984;35:95 112.
25. Dobbins WO III. Human intestinal intraepithelial lymphocytes. Gut 1986;27:972 985.
26. Bartnik W, ReMine SG, Chiba M, Thayer WR, Shorter RG. Isolation and characterization of colonic intraepithelial and lamina proprial lymphocytes. Gastroenterology 1980;78(pt 1):976 985.
27. Fawcett DW. Bloom and Fawcett: A Textbook of Histology. 11th ed. Philadelphia: WB Saunders; 1986.
28. Neutra MR, Padykula HA. The gastrointestinal tract. In: Weiss L, ed. Modern Concepts of Gastrointestinal Histology. New York: Elsevier; 1984:658 706.
29. Lewin KJ. The endocrine cells of the gastrointestinal tract. The normal endocrine cells and their hyperplasias. Part 1. In: Sommers SC, Rosen PP, Fechner RE, eds. Pathology Annual. Norwalk, CT: Appleton-Century-Croft; 1986:1 27.
30. Sjolund K, Sanden G, Hakanson R, Sundler F. Endocrine cells in human intestine: an immunocytochemical study. Gastroenterology 1983;85:1120 1130.
31. Millikin PD. Eosinophilic argentaffin cells in the human appendix. Arch Pathol 1974;98:393 395.
32. Hofler H, Kasper M, Heitz PU. The neuroendocrine system of normal human appendix, ileum and colon, and in neurogenic appendicopathy. Virchows Arch A Pathol Anat Histopathol 1983;399:127 140.
33. Sandow MJ, Whitehead R. The Paneth cell. Gut 1979;20:420 431.
34. Geller SA, Thung SN. Morphologic unity of Paneth cells. Arch Pathol Lab Med 1983;107:476 479.
35. Vestfrid MA, Suarez JE. Paneth's cells in the human appendix. A statistical study. Acta Anat (Basel) 1977;97:347 350.
36. Eastwood GL. Gastrointestinal epithelial renewal. Gastroenterology 1977;72(pt 1):962 975.
37. Lipkin M. Proliferation and differentiation of normal and diseased gastrointestinal cells. In: Johnson LR, ed. Physiology of the Gastrointestinal Tract. 2nd ed. New York: Raven Press; 1987:255 284.
38. Aubrey DA. Gastric heterotopia in the vermiform appendix. Arch Surg 1970;101:628 629.
39. Ashley DJ. Aberrant mucosa in the vermiform appendix. Br J Surg 1958;45:372 373.
40. Droga BW, Levine S, Barber JJ. Heterotopic gastric and esophageal tissue in the vermiform appendix. Am J Clin Pathol 1963;40:190 193.
P.661
41. Watson AJ. Necrosis and apoptosis in the gastrointestinal tract. Gut 1995;37:165 167.
42. Gledhill A, Cole FM. Significance of basement membrane thickening in the human colon. Gut 1984;25:1085 1088.
43. Heatley RV. The gastrointestinal mast cell. Scand J Gastroenterol 1983;18:449 453.
44. Tomasi TB Jr. Mechanisms of immune regulation at mucosal surfaces. Rev Infect Dis 1983;5(suppl 4):S784 S792.
45. Kagnoff MF. Immunology and disease of the gastrointestinal tract. In: Sleisenger M, Fordtran J, eds. Gastrointestinal Disease. 4th ed. Philadelphia: WB Saunders; 1989:114 144.
46. Spencer J, Finn T, Isaacson PG. Gut associated lymphoid tissue: a morphological and immunocytochemical study of the human appendix. Gut 1985;26:672 679.
47. van der Valk P, Meijer CJ. The histology of reactive lymph nodes. Am J Surg Pathol 1987;11:866 882.
48. Berry RJ, Lack LA. The vermiform appendix of man, and structural changes therein coincident with age. Anat Physiol 1906;40:247 256.
49. Walker NI, Bennett RE, Axelsen RA. Melanosis coli: a consequence of anthraquinone-induced apoptosis of colonic epithelial cells. Am J Pathol 1988;131:465 476.
50. Papadaki L, Rode J, Dhillon AP, Dische FE. Fine structure of a neuroendocrine complex in the mucosa of the appendix. Gastroenterology 1983;84:490 497.
51. Lundqvist M, Wilander E. Subepithelial neuroendocrine cells and carcinoid tumours of the human small intestine and appendix. A comparative immunohistochemical study with regard to serotonin, neuron-specific enolase and S-100 protein reactivity. J Pathol 1986;148:141 147.
52. Millikin PD. Extraepithelial enterochromaffin cells and Schwann cells in the human appendix. Arch Pathol Lab Med 1983;107:189 194.
53. Stanley MW, Cherwitz D, Hagen K, Snover DC. Neuromas of the appendix. A light-microscopic, immunohistochemical and electron-microscopic study of 20 cases. Am J Surg Pathol 1986;10:801 815.
54. Facer P, Bishop AE, Lloyd RV, Wilson BS, Hennessy RJ, Polak JM. Chromogranin: a newly recognized marker of endocrine cells in the human gastrointestinal tract. Gastroenterology 1985;89:1366 1373.
55. Rode J, Dhillon AP, Papadaki L. Serotonin-immunoreactive cells in the lamina propria plexus of the appendix. Hum Pathol 1983;14:464 469.
56. Sobel HJ, Marquet E, Schwarz R. Granular degeneration of appendiceal smooth muscle. Arch Pathol 1971;92:427 432.
57. Lord MG, Valies P, Broughton AC. A morphologic study of the submucosa of the large intestine. Surg Gynecol Obstet 1977;145:55 60.
58. Gershon MD, Erde SM. The nervous system of the gut. Gastroenterology 1981;80:1571 1594.
59. Krishnamurthy S, Schuffler MD. Pathology of neuromuscular disorders of the small intestine and colon. Gastroenterology 1987;93:610 639.
60. Hausman R. Granular cells in musculature of the appendix. Arch Pathol 1963;75:360 372.
61. Pieper R, Kager L, Nasman P. Clinical significance of mucosal inflammation of the vermiform appendix. Ann Surg 1983;197:368 374.
62. Petras RE. Non-neoplastic intestinal diseases. In: Mills SE, ed. Sternberg's Diagnostic Surgical Pathology, 4th ed. Philadelphia: Lippincott Williams & Wilkins; 2004:1475 1541.
63. Morson BC, Dawson IMP, Day DW, Jass JR, Price AB, Williams GT. Morson and Dawson's Gastrointestinal Pathology. 3rd ed. Oxford: Blackwell Scientific; 1990.
64. Schenken JR, Anderson TR, Coleman FC. Acute focal appendicitis. Am J Clin Pathol 1956;26:352 359.
65. Campbell JS, Fournier P, Da Silva T. When is the appendix normal? A study of acute inflammations of the appendix apparent only upon histologic examination. Can Med Assoc J 1961;85:1155 1157.
66. Touloukian RJ, Trainer TD. Significance of focal inflammation of the appendix. Surgery 1964;56:942 944.
67. Johnstone JM, Morson BC. Eosinophilic gastroenteritis. Histopathology 1978;2:335 348.
68. Klein NC, Hargrove RL, Sleisenger MH, Jeffries GH. Eosinophilic gastroenteritis. Medicine (Baltimore) 1970;49:299 319.
69. Aubock L, Ratzenhofer M. Extraepithelial enterochromaffin cell nerve-fibre complexes in the normal human appendix, and in neurogenic appendicopathy. J Pathol 1982;136:217 226.
70. Olsen BS, Holck S. Neurogenous hyperplasia leading to appendiceal obliteration: an immunohistochemical study of 237 cases. Histopathology 1987;11:843 849.
71. Qizilbash AH. Mucoceles of the appendix: their relationship to hyperplastic polyps, mucinous cystadenomas, and cystadenocarcinomas. Arch Pathol 1975;99:548 555.
72. Higa E, Rosai J, Pizzimbono CA, Wise L. Mucosal hyperplasia, mucinous cystadenoma, and mucinous cystadenocarcinoma of the appendix: a re-evaluation of appendiceal mucocele. Cancer 1973;32:1525 1541.
73. Glasser CM, Bhagavan BS. Carcinoid tumors of the appendix. Arch Pathol Lab Med 1980;104:272 275.