| Electrical Activity of Cardiac Muscle Cells In all striated muscle cells, contraction is triggered by a rapid voltage change called an action potential that occurs on the cell membrane. Cardiac muscle cell action potentials differ sharply from those of skeletal muscle cells in three important ways that promote synchronous rhythmic excitation of the heart: (1) they can be self-generating; (2) they can be conducted directly from cell to cell; and (3) they have long durations, which preclude fusion of individual twitch contractions. To understand these special electrical properties of cardiac muscle and how cardiac function depends on them, the basic electrical properties of excitable cell membranes must first be reviewed. Membrane Potentials  All cells have an electrical potential (voltage) across their membranes. Such membrane potentials exist because the ionic concentrations of the cytoplasm are different from those of the interstitium and ions diffusing down concentration gradients across semipermeable membranes generate electrical gradients. The three ions that are the most important determinants of cardiac membrane potential are sodium (Na+) and calcium (Ca2+) ions, which are more concentrated in the interstitial fluid than they are inside cells, and potassium (K+) ions, which have the opposite distribution. The diffusion of ions across the cell membrane occurs through channels that (1) are made up of protein molecules that span the membrane, (2) are specific for an individual ion (eg, Na+ channels), and (3) exist in various configurations that are open, closed, or inactivated (unable to be opened). The permeability of the membrane to a specific ion is directly related to the number of open channels for that ion at any given time. All cells have an electrical potential (voltage) across their membranes. Such membrane potentials exist because the ionic concentrations of the cytoplasm are different from those of the interstitium and ions diffusing down concentration gradients across semipermeable membranes generate electrical gradients. The three ions that are the most important determinants of cardiac membrane potential are sodium (Na+) and calcium (Ca2+) ions, which are more concentrated in the interstitial fluid than they are inside cells, and potassium (K+) ions, which have the opposite distribution. The diffusion of ions across the cell membrane occurs through channels that (1) are made up of protein molecules that span the membrane, (2) are specific for an individual ion (eg, Na+ channels), and (3) exist in various configurations that are open, closed, or inactivated (unable to be opened). The permeability of the membrane to a specific ion is directly related to the number of open channels for that ion at any given time.
Figure 2 1 shows how ion concentration differences can generate an electrical potential across the cell membrane. Consider first, as shown at the top of this figure, a cell that (1) has K+ more concentrated inside the cell than out, (2) is permeable only to K+ (ie, only K+ channels are open), and (3) has no initial transmembrane potential. Because of the concentration difference, K+ ions (positive charges) will diffuse out of the cell. Meanwhile, negative charges, such as protein anions, cannot leave the cell because the membrane is impermeable to them. Thus, the K+ efflux will make the inside of the cell more electrically negative (deficient in positively charged ions) and at the same time make the interstitium more electrically positive (rich in positive ions). Now K+ ion, being positively charged, is attracted to regions of electrical negativity. Therefore, when K+ diffuses out of a cell, it creates an electrical potential across the membrane that tends to attract it back into the cell. There exists one membrane potential called the potassium equilibrium potential at which the electrical forces tending to pull K+ into the cell exactly balance the concentration forces tending to drive K+ out. When the membrane potential has this value, there is no net movement of K+ across the membrane. With the normal concentrations of about 145 mM K+ inside cells and 4 mM K+ in the extracellular fluid, the K+ equilibrium potential is roughly 90 mV (more negative inside than outside by nine-hundredths of a volt).1 A membrane that is permeable only to K+ will inherently and rapidly (essentially instantaneously) develop the potassium equilibrium potential. In addition, membrane potential changes require the movement of so few ions that concentration differences are not significantly affected by the process. As depicted in the bottom half of Figure 2 1, similar reasoning shows how a membrane permeable only to Na+ would have the sodium equilibrium potential across it. The sodium equilibrium potential is approximately +70 mV with the normal extracellular Na+ concentration of 140 mM and intracellular concentration of 10 mM. Real cell membranes, however, are never permeable to just Na+ or just K+. When a membrane is permeable to both of these ions, the membrane potential will lie somewhere between the Na+ equilibrium potential and the K+ equilibrium potential. Just what membrane potential will exist at any instant depends on the relative permeability of the membrane to Na+ and K+. The more permeable the membrane to K+ than to Na+, the closer the membrane potential will be to 90 mV. Conversely, when the permeability to Na+ is high relative to the permeability to K+, the membrane potential will be closer to +70 mV.2 Because of low or unchanging permeabilities or low concentration, roles played by ions other than Na+ and K+ in determining membrane potential are usually minor and often ignored. However, as will be discussed later, calcium ions (Ca2+) do participate in the cardiac muscle action potential. Like Na+, Ca2+ is more concentrated outside cells than inside. The equilibrium potential for Ca2+ is  + 100 mV, and the cell membrane tends to become more positive on the inside when the membrane's permeability to Ca2+ rises. + 100 mV, and the cell membrane tends to become more positive on the inside when the membrane's permeability to Ca2+ rises. Under resting conditions, most heart muscle cells have membrane potentials that are quite close to the potassium equilibrium potential. Thus, both electrical and concentration gradients favor Na+ entry into the resting cell. However, the very low permeability of the resting membrane to Na+ in combination with an energy-requiring sodium pump that extrudes Na+ from the cell prevents Na+ from gradually accumulating inside the resting cell.3 1 The equilibrium potential (Eeq) for any ion (Xz) is determined by its intracellular and extracellular concentrations as indicated in the Nernst equation: 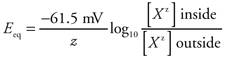
2 A quantitative description of how Na+ and K+ concentrations and the relative permeability (PNa/PK) to these ions affect membrane potential (Em) is given by the following equation: 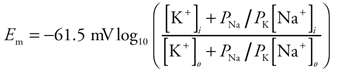
3 The sodium pump not only removes Na+ from the cell but also pumps K+ into the cell. Since more Na+ is pumped out than K+ is pumped in (3:2), the pump is said to be electrogenic. The resting membrane potential becomes slightly less negative than normal when the pump is abruptly inhibited. Cardiac Cell Action Potentials  Action potentials of cells from different regions of the heart are not identical but have varying characteristics that are important to the overall process of cardiac excitation. Action potentials of cells from different regions of the heart are not identical but have varying characteristics that are important to the overall process of cardiac excitation.
 Some cells within a specialized conduction system have the ability to act as pacemakers and to spontaneously initiate action potentials whereas ordinary cardiac muscle cells do not (except under unusual conditions). Basic membrane electrical features of an ordinary cardiac muscle cell and a cardiac pacemaker-type cell are shown in Figure 2 2. Action potentials from these cell types are referred to as "fast response" and "slow response" action potentials, respectively. As shown in panel A of this figure, fast response action potentials are characterized by a rapid depolarization (phase 0) with a substantial overshoot (positive inside voltage), a rapid reversal of the overshoot potential (phase 1), a long plateau (phase 2), and a repolarization (phase 3) to a stable, high (ie, large negative) resting membrane potential (phase 4). In comparison, the slow response action potentials are characterized by a slower initial depolarization phase, a lower amplitude overshoot, a shorter and less stable plateau phase, and a repolarization to an unstable, slowly depolarizing "resting" potential (Figure 2 2B). The unstable resting potential seen in pacemaker cells with slow response action potentials is variously referred to as the phase 4 depolarization, diastolic depolarization, or pacemaker potential. Some cells within a specialized conduction system have the ability to act as pacemakers and to spontaneously initiate action potentials whereas ordinary cardiac muscle cells do not (except under unusual conditions). Basic membrane electrical features of an ordinary cardiac muscle cell and a cardiac pacemaker-type cell are shown in Figure 2 2. Action potentials from these cell types are referred to as "fast response" and "slow response" action potentials, respectively. As shown in panel A of this figure, fast response action potentials are characterized by a rapid depolarization (phase 0) with a substantial overshoot (positive inside voltage), a rapid reversal of the overshoot potential (phase 1), a long plateau (phase 2), and a repolarization (phase 3) to a stable, high (ie, large negative) resting membrane potential (phase 4). In comparison, the slow response action potentials are characterized by a slower initial depolarization phase, a lower amplitude overshoot, a shorter and less stable plateau phase, and a repolarization to an unstable, slowly depolarizing "resting" potential (Figure 2 2B). The unstable resting potential seen in pacemaker cells with slow response action potentials is variously referred to as the phase 4 depolarization, diastolic depolarization, or pacemaker potential.
As indicated at the bottom of Figure 2 2A, cells are in an absolute refractory state during most of the action potential (ie, they cannot be stimulated to fire another action potential). Near the end of the action potential, the membrane is relatively refractory and can be reexcited only by a larger than normal stimulus. Immediately after the action potential, the membrane is transiently hyperexcitable and is said to be in a "vulnerable" or "supranormal" period. Similar alterations in membrane excitability probably occur during slow action potentials but at present are not fully characterized.  Recall that the membrane potential of any cell depends on the relative permeability of the cell's membrane to specific ions at that instant. As in all excitable cells, cardiac cell action potentials are the result of transient changes in the ionic permeability of the cell membrane, which are triggered by an initial depolarization. Panels C and D of Figure 2 2 indicate the changes in membrane permeabilities to K+, Na+, and Ca2+, which produce the various phases of the fast and slow response action potentials. Note that during the resting phase, the membranes of both types of cells are more permeable to K+ than to Na+ or Ca2+. Therefore, the membrane potentials are close to the potassium equilibrium potential (of 90 mV) during this period. In the pacemaker-type cells, at least three mechanisms are thought to contribute to the slow depolarization of the membrane observed during the diastolic interval. First, there is a progressive decrease in the membrane s permeability to K+ during the resting phase, and second, the permeability to Na+ increases slightly. The gradual increase in the Na+/K+ permeability ratio will cause the membrane potential to move slowly away from the K+ equilibrium potential ( 90 mV) in the direction of the Na+ equilibrium potential. Third, there is an increase in the permeability of the membrane to calcium ions, which results in an inward movement of positively charged ions and also contributes to the diastolic depolarization. Recall that the membrane potential of any cell depends on the relative permeability of the cell's membrane to specific ions at that instant. As in all excitable cells, cardiac cell action potentials are the result of transient changes in the ionic permeability of the cell membrane, which are triggered by an initial depolarization. Panels C and D of Figure 2 2 indicate the changes in membrane permeabilities to K+, Na+, and Ca2+, which produce the various phases of the fast and slow response action potentials. Note that during the resting phase, the membranes of both types of cells are more permeable to K+ than to Na+ or Ca2+. Therefore, the membrane potentials are close to the potassium equilibrium potential (of 90 mV) during this period. In the pacemaker-type cells, at least three mechanisms are thought to contribute to the slow depolarization of the membrane observed during the diastolic interval. First, there is a progressive decrease in the membrane s permeability to K+ during the resting phase, and second, the permeability to Na+ increases slightly. The gradual increase in the Na+/K+ permeability ratio will cause the membrane potential to move slowly away from the K+ equilibrium potential ( 90 mV) in the direction of the Na+ equilibrium potential. Third, there is an increase in the permeability of the membrane to calcium ions, which results in an inward movement of positively charged ions and also contributes to the diastolic depolarization.
When the membrane potential depolarizes to a certain threshold potential in either type of cell, major rapid alterations in the permeability of the membrane to specific ions are triggered. Once initiated, these permeability changes cannot be stopped and they proceed to completion. The characteristic rapid rising phase of the fast response action potential is a result of a sudden increase in Na+ permeability. This produces what is referred to as the fast inward current of Na+ and causes the membrane potential to move rapidly toward the sodium equilibrium potential. As indicated in panel C of Figure 2 2, this period of very high sodium permeability is short-lived. It is followed by a more slowly developed increase in the membrane's permeability to Ca2+ and a decrease in its permeability to K+. Also, there is a second slowly developing increase in Na+ permeability, which is thought to be caused by a different mechanism than that involved in the initial rapid Na+ permeability changes. These more persistent permeability changes (which produce what is referred to as the slow inward current) prolong the depolarized state of the membrane to cause the plateau (phase 2) of the cardiac action potential. The initial fast inward current is small (or even absent) in cells that have slow response action potentials. The slow rising phase of these action potentials is therefore primarily a result of an inward movement of Ca2+ ions. In both types of cells, the membrane is repolarized (phase 3) to its original resting potential as the K+ permeability increases and the Ca2+ and Na+ permeabilities return to their low resting values. These late permeability changes produce what is referred to as the delayed outward current. The overall smoothly graded permeability changes that produce action potentials are the net result of alterations in each of the many individual ion channels within the plasma membrane of a single cell. The experimental technique of patch clamping has made it possible to study the operation of individual ion channels. The patch clamp data clearly indicate that a single channel is either open or closed at any instant in time; there are no graded states of partial opening. What is graded is the percentage of time that a channel spends in the open state, ie, its probability of being open. While a channel may remain closed for long periods, it rarely remains open for more than a few milliseconds at a time. Thus, the probability of a channel's being open depends both on the frequency with which it opens and how long it remains open. An increase in an ion channel's probability of being open (channel "activation") leads to an increase in total open time and an increase in the overall membrane permeability to that ion. Certain types of channels are called voltage-gated channels (or voltage-operated channels) because their probability of being open varies with membrane potential. Other types of channels, called ligand-gated channels (or receptor-operated channels), are activated by certain neurotransmitters or other specific signal molecules. Table 2 1, at the end of this section, lists some of the major currents and channel types involved in cardiac cell electrical activity. | Table 2 1. Characteristics of Important Cardiac Ion Channels in Order of Their Participation in an Action Potential.
| | | Current | Channel | Gating mechanism | Functional role | iK1
| K+ channel (inward rectifier)
| Voltage | Maintains high K+ permeability during phase 4 Its decay contributes to diastolic depolarization Its suppression during phases 0 to 2 contribute to plateau | iNa
| Na+ channel (fast)
| Voltage | Accounts for phase 0 of action potential Inactivation may contribute to phase 1 of action potential | iTo
| K+ channel (transient outward)
| Voltage | Contributes to phase 1 of action potential | iCa
| Ca2+ channel (slow inward, L channels)
| Voltage | Primarily responsible for phase 2 of action potential Inactivation may contribute to phase 3 of action potential Is enhanced by sympathetic stimulation and 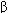 -adrenergic agents -adrenergic agents | iK
| K+ channel (delayed rectifier)
| Voltage | Causes phase 3 of action potential May be enhanced by increased intracellular Ca2+ | iKATP
| K+ channel (ATP-sensitive)
| Ligand | Increases K+ permeability when [ATP] is low
| iKACh
| K+ channel (acetylcholine-activated)
| Ligand | Responsible for effects of vagal stimulation Decreases diastolic depolarization (and heart rate) Hyperpolarizes resting membrane potential Shortens phase 2 of the action potential | if("funny")
| Na+ channel (pacemaker current)
| Both | Contributes to the diastolic depolarization Is enhanced by sympathetic stimulation and -adrenergic agents Is suppressed by vagal stimulation | |
| Some of the voltage-gated channels respond to a sudden onset, sustained change in membrane potential by only a brief period of activation. However, changes in membrane potential of slower onset but the same magnitude may fail to activate these channels at all. To explain such behavior, it is postulated that these channels have two independently operating "gates" an activation gate and an inactivation gate both of which must be open for the channel as a whole to be open. These gates both respond to changes in membrane potential but do so with different voltage sensitivities and time courses. These concepts are illustrated in Figure 2 3. In the resting state, with the membrane polarized to near 80 mV, the activation or m gate of the fast Na+ channel is closed, but its inactivation or h gate is open (Figure 2 3A). With a rapid depolarization of the membrane to threshold, the Na+ channels will be activated strongly to allow an inrush of positive sodium ions that further depolarizes the membrane and thus initiates a "fast" response action potential as illustrated in Figure 2 3B. This occurs because the m gate responds to membrane depolarization by opening more quickly than the h gate responds by closing. Thus a rapid depolarization to threshold is followed by a brief but strong period of Na+ channel activation wherein the m gate is open but the h gate has yet to close.  The initial membrane depolarization also causes the activation (d) gate of the Ca2+ channel to open after a brief delay. This permits the slow inward current of Ca2+ ions, which helps maintain the depolarization through the plateau phase of the action potential (Figure 2 3C). Ultimately, repolarization occurs because of both a delayed inactivation of the Ca2+ channel (by closure of the f gates) and an opening of K+ channels (which are not shown in Figure 2 3). Multiple factors influence the operation of K+ channels. For example, high intracellular Ca2+ concentration contributes to activation of K+ channels during repolarization. The h gates of sodium channels remain closed during the remainder of the action potential, effectively inactivating the Na+ channel and contributing to the long cardiac refractory period which lasts until the end of phase 3. With repolarization, both gates of the sodium channel return to their original position and the channel is now ready to be reactivated by a subsequent depolarization. The initial membrane depolarization also causes the activation (d) gate of the Ca2+ channel to open after a brief delay. This permits the slow inward current of Ca2+ ions, which helps maintain the depolarization through the plateau phase of the action potential (Figure 2 3C). Ultimately, repolarization occurs because of both a delayed inactivation of the Ca2+ channel (by closure of the f gates) and an opening of K+ channels (which are not shown in Figure 2 3). Multiple factors influence the operation of K+ channels. For example, high intracellular Ca2+ concentration contributes to activation of K+ channels during repolarization. The h gates of sodium channels remain closed during the remainder of the action potential, effectively inactivating the Na+ channel and contributing to the long cardiac refractory period which lasts until the end of phase 3. With repolarization, both gates of the sodium channel return to their original position and the channel is now ready to be reactivated by a subsequent depolarization.
The slow response action potential shown in the right half of Figure 2 3 differs from the fast response action potential primarily because of the lack of a strong activation of the fast Na+ channel at its onset. This is a direct consequence of a slow depolarization to the threshold potential. Slow depolarization gives the inactivating h gates time to close even as the activating m gates are opening (Figure 2 3D). Thus, in a slow response action potential, there is no initial period where all the sodium channels of a cell are essentially open at once. The depolarization beyond threshold is slow and caused primarily by the influx of Ca2+ through slow channels (Figure 2 3E). While cells in certain areas of the heart typically have fast-type action potentials and cells in other areas normally have slow-type action potentials, it is important to recognize that all cardiac cells are potentially capable of having either type of action potential depending on how fast they depolarize to the threshold potential. As we shall see, rapid depolarization to the threshold potential is usually an event forced on a cell by the occurrence of an action potential in an adjacent cell. Slow depolarization to threshold occurs when a cell itself spontaneously and gradually loses its resting polarization, which normally happens only in the sinoatrial (SA) node. A chronic moderate depolarization of the resting membrane (caused, for example, by moderately high extracellular K+ concentration) can inactivate the fast channels (by closing the h gates) without inactivating the slow Ca2+ channels. Under these conditions, all cardiac cell action potentials will be of the slow type. Large sustained depolarizations, however, can inactivate both the fast and slow channels and thus make the cardiac muscle cells inexcitable. Conduction of Cardiac Action Potentials  Action potentials are conducted over the surface of individual cells because active depolarization in any one area of the membrane produces local currents in the intracellular and extracellular fluids which passively depolarize immediately adjacent areas of the membrane to their voltage threshold for active depolarization. Action potentials are conducted over the surface of individual cells because active depolarization in any one area of the membrane produces local currents in the intracellular and extracellular fluids which passively depolarize immediately adjacent areas of the membrane to their voltage threshold for active depolarization.
In the heart, cardiac muscle cells are connected end-to-end by structures called intercalated disks. These disks contain the following: (1) firm mechanical attachments between adjacent cell membranes by proteins called adherins in structures called desmosomes and (2) low resistance electrical connections between adjacent cells through channels formed by protein called connexin in structures called gap junctions. Figure 2 4 shows schematically how these gap junctions allow action potential propagation from cell to cell.
Cells B, C, and D are shown in the resting phase with more negative charges on the inside than the outside. Cell A is shown in the plateau phase of an action potential and has more positive charges inside than out. Because of the gap junctions, electrostatic attraction can cause a local current flow (ion movement) between the depolarized membrane of active cell A and the polarized membrane of resting cell B, as indicated by the arrows in the figure. This ion movement tends to eliminate the charge difference across the resting membrane; ie, it depolarizes the membrane of cell B. Once the local currents from active cell A depolarize the membrane of cell B near the gap junction to the threshold level, an action potential will be triggered at that site and will be conducted over cell B. Because cell B branches (a common morphological characteristic of cardiac muscle fibers), its action potential will evoke action potentials on cells C and D. This process is continued through the entire myocardium. Thus, an action potential initiated at any site in the myocardium will be conducted from cell-to-cell throughout the entire myocardium. The speed at which an action potential propagates through a region of cardiac tissue is called the conduction velocity. The conduction velocity varies considerably in different areas in the heart. This velocity is directly dependent on the diameter of the muscle fiber involved. Thus, conduction over small-diameter cells in the atrioventricular (AV) node is significantly slower than conduction over large-diameter cells in the ventricular Purkinje system. Conduction velocity is also directly dependent on the intensity of the local depolarizing currents, which are in turn directly determined by the rate of rise of the action potential. Rapid depolarization favors rapid conduction. Variations in the capacitive and/or resistive properties of the cell membranes, gap junctions, and cytoplasm are also factors that contribute to the differences in conduction velocity of action potentials through specific areas of the heart. Details of the cardiac conduction system are shown in Figure 2 5. Specific electrical adaptations of various cells in the heart are reflected in the characteristic shape of their action potentials, as shown in the right half of Figure 2 5. Note that the action potentials shown in Figure 2 5 have been positioned to indicate the time at which the electrical impulse that originates in the SA node reaches other areas of the heart. Cells of the SA node act as the heart's normal pacemaker and determine the heart rate. This is because the spontaneous depolarization of the resting membrane is most rapid in SA nodal cells, and they reach their threshold potential before cells elsewhere. The action potential initiated by an SA nodal cell first spreads progressively through the atrial wall. Action potentials from cells in two different regions of the atria are shown in Figure 2 5: one close to the SA node and one more distant from the SA node. Both cells have similarly shaped action potentials, but their temporal displacement reflects the fact that it takes some time for the impulse to spread over the atria. As shown in Figure 2 5, action potential conduction is greatly slowed as it passes through the AV node. This is because of the small size of the AV nodal cells and the slow rate of rise of their action potentials. Since the AV node delays the transfer of the cardiac excitation from the atria to the ventricles, atrial contraction can contribute to ventricular filling just before the ventricles contract. Note also that AV nodal cells have a faster spontaneous depolarization during the resting period than other cells of the heart except those of the SA node. The AV node is sometimes referred to as a latent pacemaker, and in many pathological situations it (rather than the SA node) controls the heart rhythm. Because of sharply rising action potentials and other factors, such as large cell diameters, electrical conduction is extremely rapid in Purkinje fibers. This allows the Purkinje system to transfer the cardiac impulse to cells in many areas of the ventricle nearly in unison. Action potentials from muscle cells in two areas of the ventricle are shown in Figure 2 5. Because of the high conduction velocity in ventricular tissue, there is only a small discrepancy in their time of onset. Note in Figure 2 5 that the ventricular cells which are the last to depolarize have shorter duration action potentials and thus are the first to repolarize. The physiological importance of this unexpected behavior is not clear but it does have an influence on the electrocardiograms that will be discussed in Chapter 4. Electrocardiograms Fields of electrical potential caused by the electrical activity of the heart extend through the body tissue and may be measured with electrodes placed on the body surface. Electrocardiography provides a record of how the voltage between two points on the body surface changes with time as a result of the electrical events of the cardiac cycle. At any instant of the cardiac cycle the electrocardiogram indicates the net electrical field that is the summation of many weak electrical fields being produced by voltage changes occurring on individual cardiac cells at that instant. When a large number of cells are simultaneously depolarizing or repolarizing, large voltages are observed on the electrocardiogram. Since the electrical impulse spreads through the heart tissue in a stereotyped manner, the temporal pattern of voltage change recorded between two points on the body surface is also stereotyped and will repeat itself with each heart cycle. The lower trace of Figure 2 5 represents a typical recording of the voltage changes normally measured between the right arm and the left leg as the heart goes through two cycles of electrical excitation; this record is called a lead II electrocardiogram and will be discussed in detail in Chapter 4. The major features of an electrocardiogram are the P wave, the QRS complex, and the T wave. The P wave corresponds to atrial depolarization, the QRS complex to ventricular depolarization, and the T wave to ventricular repolarization. Control of Heart Beating Rate Normal rhythmic contractions of the heart occur because of spontaneous electrical pacemaker activity (automaticity) of cells in the SA node. The interval between heartbeats (and thus the heart rate) is determined by how long it takes the membranes of these pacemaker cells to spontaneously depolarize to the threshold level. The SA nodal cells fire at a spontaneous or intrinsic rate (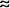 100 beats per minute) in the absence of any outside influences. Outside influences are required, however, to increase or decrease automaticity from its intrinsic level. 100 beats per minute) in the absence of any outside influences. Outside influences are required, however, to increase or decrease automaticity from its intrinsic level.  The two most important outside influences on automaticity of SA nodal cells come from the autonomic nervous system. Fibers from both the sympathetic and parasympathetic divisions of the autonomic system terminate on cells in the SA node and these fibers can modify the intrinsic heart rate. Activating the cardiac sympathetic nerves (increasing cardiac sympathetic tone) increases the heart rate. Increasing cardiac parasympathetic tone slows the heart. As shown in Figure 2 6, the parasympathetic and sympathetic nerves both influence heart rate by altering the course of spontaneous depolarization of the resting potential in SA pacemaker cells. The two most important outside influences on automaticity of SA nodal cells come from the autonomic nervous system. Fibers from both the sympathetic and parasympathetic divisions of the autonomic system terminate on cells in the SA node and these fibers can modify the intrinsic heart rate. Activating the cardiac sympathetic nerves (increasing cardiac sympathetic tone) increases the heart rate. Increasing cardiac parasympathetic tone slows the heart. As shown in Figure 2 6, the parasympathetic and sympathetic nerves both influence heart rate by altering the course of spontaneous depolarization of the resting potential in SA pacemaker cells.
Cardiac parasympathetic fibers, which travel to the heart through the vagus nerves, release the transmitter substance acetylcholine on SA nodal cells. Acetylcholine increases the permeability of the resting membrane to K+ and decreases the diastolic permeability to Na+.4 As indicated in Figure 2 6, these permeability changes have two effects on the resting potential of cardiac pacemaker cells: (1) they cause an initial hyperpolarization of the resting membrane potential by bringing it closer to the K+ equilibrium potential and (2) they slow the rate of spontaneous depolarization of the resting membrane. Both these effects increase the time between beats by prolonging the time required for the resting membrane to depolarize to the threshold level. Since there is normally some continuous tonic activity of cardiac parasympathetic nerves, the normal resting heart rate is approximately 70 beats per minute. Sympathetic nerves release the transmitter substance norepinephrine on cardiac cells. In addition to other effects discussed later, norepinephrine increases the inward currents carried by Na+ (if) and by Ca2+ during the diastolic interval.5 These changes will increase heart rate by increasing the rate of diastolic depolarization as shown in Figure 2 6. In addition to sympathetic and parasympathetic nerves, there are many (usually less important) factors that can alter heart rate. These include a number of ions and circulating hormones, as well as physical influences such as temperature and atrial wall stretch. All act by somehow altering the time required for the resting membrane to depolarize to the threshold potential. An abnormally high concentration of Ca2+ in the extracellular fluid, for example, tends to decrease heart rate by shifting the threshold potential. Factors that increase heart rate are said to have a positive chronotropic effect. Those that decrease heart rate have a negative chronotropic effect. Besides their effect on heart rate, autonomic fibers also influence the conduction velocity of action potentials through the heart. Increases in sympathetic activity increase conduction velocity (have a positive dromotropic effect), whereas increases in parasympathetic activity decrease conduction velocity (have a negative dromotropic effect). These effects are most notable at the AV node and can influence the duration of the PR interval. 4 Acetylcholine interacts with a muscarinic receptor on the SA nodal cell membrane which in turn is linked to an inhibitory G protein, Gi. The activation of Gi has two effects: (1) an increase in K+ conductance resulting from an increased opening of the KAch channels and (2) a suppression of adenylate cyclase leading to a fall in intracellular cyclic adenosine monophosphate (cAMP) which reduces the inward-going pacemaker current carried by Na+ (if). 5 Norepinephrine interacts with 1-adrenergic receptors on the SA nodal cell membrane which in turn are linked to stimulatory G proteins, Gs. The activation of Gs increases adenylate cyclase, leading to an increase in intracellular cyclic AMP which increases the open-state probability of the pacemaker Na+ current channel (if). |