1 - Normal Skin
Editors: Mills, Stacey E.
Title: Histology for Pathologists, 3rd Edition
Copyright 2007 Lippincott Williams & Wilkins
> Table of Contents > I - Cutaneous Tissue > 2 - Nail
function show_scrollbar() {}
2
Nail
Julian Conejo-Mir
Luis Requena
Introduction
Fingernails are an important epithelial mini-organ system of the hand, with a complex anatomical structure well described in the last decades. They are important in certain animals for the apprehension and capture of prey. In primates and humans, the nails have two different functions: as a protective plate and to enhance sensation of the fingertip. The protection function of the fingernail is commonly known, but the sensation function is equally important. The fingertip has many nerve endings in it allowing us to receive volumes of information about objects we touch. The nail acts as a counterforce to the fingertip providing even more sensory input when an object is touched.
Although most pathological specimens from the nail show well-known changes such as psoriasis, lichen planus, and the characteristic malignant tumors, a broad spectrum of other changes may be found. For the pathologist, knowledge of the normal histology and its more common variations is important in establishing a correct diagnosis. Unfortunately, much of the literature on the nail can be troublesome and confusing because a great profusion of names and concepts exists and has changed over the years; also, many newer concepts of the embryology, physiology, genetics, immunohistochemistry, and nail growth mechanism find their way slowly into textbooks. This chapter emphasizes those observations and theories related to clinical pathology.
History
Historical interest can be traced to the works of Galen in the second century BC when he noticed the nail's resemblance to hair structure. However, the real study of the nail begins at the end of the nineteenth century, mostly by Germans like Zander (1), Kolliker (2), and Unna (3). The first studies dealt with embryology and anatomy and their comparison to birds and primates (1,4,5). After the initial spark of interest, the nail literature was enriched by many authors,
P.30
both on the embryology and the anatomy of the human nail (6,7). Because of the technical shortcomings of their time, the authors interpreted the nail plate as formed entirely by the matrix cells and concluded that other adjacent structures did not contribute in the formation of the plate.
During the 1950s, Lewis (8) challenged this view and published his idea of the nail unit, consisting of a dorsal, intermediate, and ventral nail, with differentiation based on the use of a silver-proteinate stain. In 1963, Zaias (9) extended the concept of the nail unit, including the proximal nail fold, the matrix, the nail bed, and the hyponychium, all of which contribute to the formation of the nail.
During the last 25 years, most studies on the nail have fundamentally tried to explain its biochemistry and physiology, with emphasis on analyzing nail growth; also considered were ultrastructure and, most recently, the immunohistochemistry of the nail. At the same time, new conditions, particularly tumors, unknown until a few years ago, have been described from the point of ungual structures, especially keratoacanthomas, merkelomas (Merkel cell tumors), and subungual Bowen's disease. In the last five years, however, publications have been very scarce following the great proliferation of papers in previous years.
Difficult biopsy access, as well as the complicated orientation and specimen handling (with the resulting difficulties of interpretation), are the main reasons why there are few histologic and histopathologic studies on the nail.
Embryology
Whereas the embryonic development of the fetal skin has been divided into eight stages, using scanning electron microscopy (10), the embryonic development of the nail shows only five stages (11,12): (a) plate phase; (b) fibrillar phase; (c) granular phase; (d) squamous phase; and (e) definitive nail phase, or end phase (Table 2.1).
The earliest recognizable fingers are seen in 42- to 45-day-old embryos (16 mm crown-rump), while the toes lag somewhat and are seen at 52 to 54 days of age (18.5 mm) (13). Studies using optical microscopy showed that the ungual morphogenesis begins at the embryonal age of 10 weeks, with a smooth, shiny quadrangular surface delineated by continuous shallow grooves. This surface of the phalanx is the primary nail base of Zander (1) or primary nail field of Zaias (9), delimited proximally by a transversal groove (the proximal nail groove).
Studies we performed using scanning electron microscopy showed that the formation of the nail begins very early, at the embryonal age of seven weeks, with an accumulation of strongly active cells, abundant mitosis, and cellular damage, followed by necrosis, with the presence of T lymphocytes in the primary nail base (Figure 2.1). This phenomenon, named apoptosis, occurs in all the epidermal accumulated cells following a transversal band in the dorsal area of the distal third of the fingers. Apoptosis of these epidermal cells is the most important step in the nail's development because it permits an immediate epidermal invagination identical to the one in the hair follicle except for one difference: in the hair follicle, the process starts at the age of 2.5 to 3 months. We observed apoptosis in nail development in this first phase only. Yet, the two processes are so identical that sometimes the layers of the nail have been compared with those of the hair follicle. The result is the formation of a transversal groove, which subsequently becomes the proximal nail fold.
Table 2.1 Comparison of the Different Stages of Nail Development with the Epidermis of the Embryo Using Scanning Electron Microscopy | |||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||
An interesting feature of the first stages is the excessive size of the primitive nail plate (2.5 to 3 months), nearly occupying the total distal third part of the finger. This plate stays attached to its surroundings through some periungual-fixing filaments (Figure 2.2). At the age of 11 weeks, all the proximal and lateral nail folds are already formed. The transversal distal fold, corresponding to the hyponychium, is completely keratinized at the age of 3.5 months (Figure 2.3). Afterward, the epidermal cells of the nail field suffer a process of keratin formation that is different from the rest of the embryo. The result is a keratinized structure, covering the whole nail bed from the age of 14 weeks on and sometimes confused by some authors with a false nail (Figure 2.4) (8,9). The production of the true nail plate starts from the matrix cells, located in the proximal nail groove and the most proximal portion of the nail bed. Its presence in the
P.31
proximal fold is visible from the fifth month of intrauterine life onward, the histochemical confirmation of its formation being the presence of sulfhydryl radicals (14).
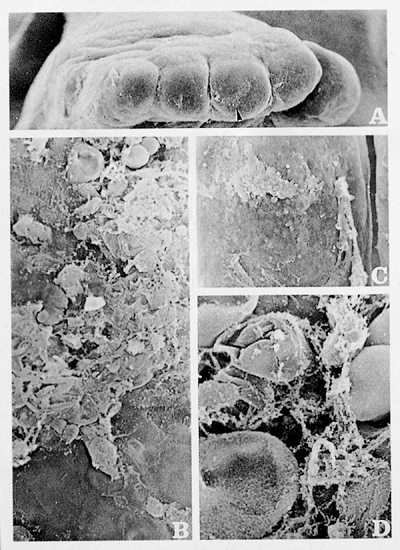 |
Figure 2.1 Development of the human nail exhibited through scanning electron microscopy (13). A. Plaque phase: foot of 7-week-old human embryo. The fingers are already defined but have no interphalangeal folds ( 50). Distally on the third toe, you can see poorly structured material accumulated (arrow in A) that corresponds to apoptotic cells. B and C. These apoptotic cells limit the future proximal nail fold (B, 500; C, 100). D. Close-up view of the apoptotic cells: amorphous extracellular material appears with numerous vesicles of keratohyalin, which are in different phases of their evolution ( 1500). |
The nail unit at this stage shows grooves form by invaginations of primitive ectoderm in regions that will become nail folds. These grooves delimit rectangular areas at distal aspects of dorsa of fingers and toes where nail plates will be situated subsequently. These areas are covered by primitive epithelium that, approximately at the fourteenth week of intrauterine-life (Figure 2.5), appears composed of a basal layer of primitive germinative cells, three or four layers of primitive keratinocytes with clear or pale cytoplasm, and a thin and eosinophilic acellular layer at the surface. This primitive epithelium covering the dorsum of a distal phalanx develops two clusters of epithelial cells at their proximal and distal ends. The proximal bud of primitive epithelial basaloid cells proliferates backward and downward, descending at an angle into the subjacent primitive mesenchymal tissue. The superficial part of this wedge of epithelial cells will become the proximal nail fold, and the deeper part will eventuate in the dorsal and intermediate nail matrices. At the junction between the superficial and deeper parts, there is a crease of cornified cells that will be the cuticle of the fully developed nail. At this stage of development, the primitive mesenchymal tissue underlying the future nail is a highly cellular tissue with abundant ground substance. At this time, the future distal phalanx is represented by primitive cartilaginous tissue with the earliest evidences of focal calcification. Distally, the primitive epithelium forms another cluster of cells with a distal ridge that will become the hyponychium.
From the fifth month on, the definitive nail plate starts to grow in a distal sense until it reaches the hyponychium at the time of birth. The growth mechanism of the definitive nail is discussed later in the section entitled Nail Growth.
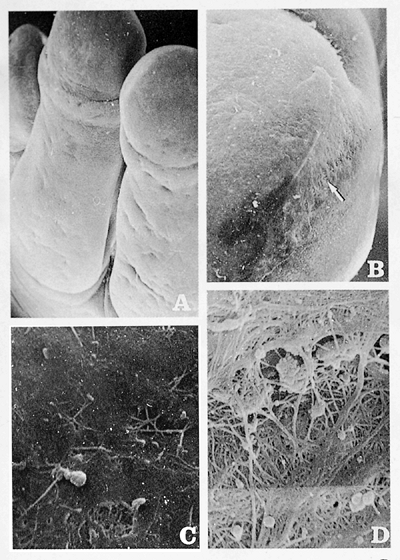 |
Figure 2.2 Fibrillar phase. Fingers of the hand of a 3-month-old embryo. A. The ungual region is perfectly delimited by the proximal nail fold ( 40). B. The ungual region is delimited by multiple fibrillar formations ( 150). C. Different morphology is seen in the nail bed surface ( 2400). D. Detail of the fibrillar attachment of the nail region to the neighboring tissue ( 2400). |
P.32
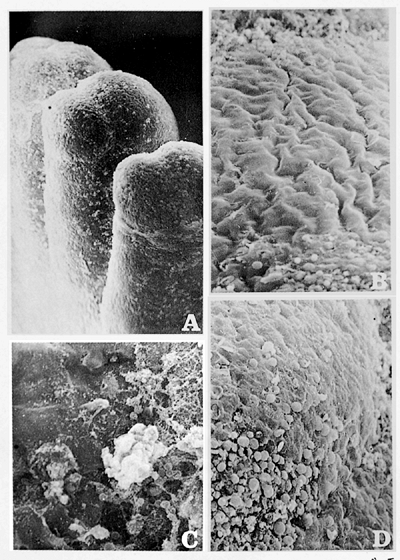 |
Figure 2.3 Granular phase. Fingers of the hand of 4.5-month-old embryo. A. All fingers show a granular aspect ( 40). B. The nail bed has an undulating surface covered by keratin scales ( 400). C. and D. The hyponychium zone is occupied by numerous keratohyalin vesicles (C, 400; D, 150). |
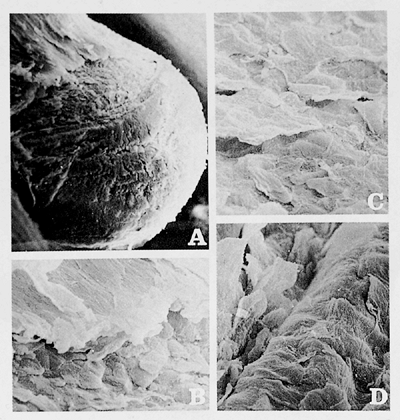 |
Figure 2.4 Squamous phase A. Index finger of 5.5-month-old fetus ( 200). C. The keratinization process is complete in the nail bed surface, simulating a false nail ( 500). B. The cuticle ( 500) and D. The hyponychium are also completely developed ( 500). |
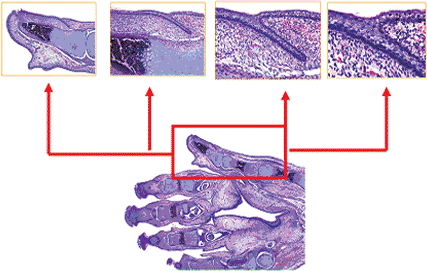 |
Figure 2.5 Sagittal section of a fetus hand of 16 weeks, with special close up views of the nail matrix area. |
P.33
Genetics and Nail Keratins
Epithelial cells possess three cytoskeletal systems: actin microfilaments, microtubules, and keratin intermediate filaments. The protective structural role of keratins was clearly revealed in the early 1990s, when mutations in human keratin genes were discovered in a variety of human genetic diseases characterized by fragility and/or overgrowth (hyperkeratosis) of specific epithelial tissues (15). It is not precisely known how mutations in these keratins lead to hyperkeratosis of the nail, but fragility of the underlying nail bed keratinocytes presumably leads to release of cytokines and other inflammatory factors that act upon the proliferative cells of the nail matrix and produce overgrowth of the nail (16).
Keratins are a large family of intermediate filament proteins encoded by more than 50 distinct genes in humans (17). About half of these are the epithelial keratins that are found in soft epithelial tissues of the human body. The rest are the trichocyte or high-sulphur hard keratins of which hair and nail is composed. Both epithelial keratins and hard keratins can be further subdivided into type I and type II proteins on the basis of their size, charge, and amino acid sequence characteristics (18). Recent work has shown that the human hair keratin family consists of nine type I and six type II members, whose genes are organized as distinct clusters within the type I and type II epithelial keratin gene domains on chromosomes 17q21.2 and 12q13.3, respectively (16). The functional type I (K9 K23; Ha1 Ha8) and type II keratin genes (K1 K8; Hb1 Hb6) are each clustered on distinct chromosomes in the human and mouse genomes. The pairwise and differentiation-related regulation of most type I and type II keratin genes provides a unique handle to track differentiation within epithelial tissue (19). A family-wide, crucial function of keratin filaments is to endow epithelial cells with the ability to withstand mechanical and other forms of stress.
Accordingly, mutations in keratin genes are responsible for a number of genetically based fragility disorders involving specific cell types in skin and other epithelia (20,21). Among type I keratin genes, K17 stands out in multiple ways. In mouse embryonic skin, it is first expressed in ectodermal cells committing to a nonepidermal cell fate (i.e., all appendages and periderm) in response to mesenchymal induction (22). Concomitant with skin maturation, mK17 expression becomes restricted to specific cell layers and compartments within all major types of epithelial appendages. Both hK17 and mK17 can be coregulated with distinct type II keratin genes (e.g., K5, K6a, K6b, K6hf) in mature epithelial settings (23). In addition to its constitutive expression in epithelial appendages, K17 expression is induced in mature interfollicular epidermis subjected to various types of acute challenges (e.g., injury, ultraviolet light exposure, inflammation) or during diseases (e.g., psoriasis, basal cell carcinoma). Mutations affecting a particular segment of hK17's coding sequence can cause distinct disorders of the skin, related to ectodermal dysplasias (15).
Several genodermatoses, such as Darier's disease, X-linked dyskeratosis congenita (DC), and pachyonychia congenita (PC), are associated with characteristic nail changes (24). A causative gene for Darier's disease, which resides at 12q23-q24.1, encodes the sarco/endoplasmic reticulum Ca2+-ATPase type 2 isoform. Mutations in XAP101 gene, which maps to Xq28, are responsible for DC. Thus, mutations in DKC1 gene encoding dyskerin have been also found (25). Pachyonychia congenita is a group of autosomal dominant ectodermal dysplasias, whose most obvious phenotype is hypertrophic nail dystrophy leading to a phenotype of grossly thickened nail. There are two main types of PC: the Jadassohn Lewandowsky form (type 1) and the Jackson Lawler form (type 2). The epithelia affected in pachyonychia congenita type 1 express the keratin pair K6a and K16, and the condition is caused by dominant-negative mutations in these genes (26). The pachyonychia congenita type 2 variant is caused by mutations in keratins K6b and K17 (27). All the tissues affected in pachyonychia congenita type 2 express K6b and K17 (28,29,30).
Gross Anatomy
It was first noted early in the twentieth century that the nail unit was comparable in several respects to a hair follicle sectioned longitudinally and laid on its side. The epithelial components of the hair follicle and nail apparatus are differentiated epidermal structures that may be involved jointly in several ways as congenital and hereditary anomalies and acquired conditions such as alopecia areata, lichen planus, iatrogenic causes, and fungal infection (31).
Various types of differently keratinizing epidermis make up the nail. What commonly is termed the nail plate is the horny end product of the most important epidermal component, the matrix. Usually, this nail plate is slightly convex or flat, rectangular, and of varying size between approximately 1 1 and 2 3 cm, depending on the finger (Figures 2.6,2.7,2.8). In the hand, this is usually 25 to 50% of the dorsal surface of the fingertips, whereas in the big toe it occupies about 75%. The nail is translucent and becomes rosy from the underlying vascular network. However, change of colors (erythronychia, melanonychia) can be observed in the nail plate and may indicate inflammatory diseases (lichen planus, lupus erythematosus); benign or malignant neoplasms, main subungual melanoma; and scarring of the dermis or epidermis (32). The white appearance of nails in leukonychia seems to be a result of an abnormal keratinization of cells originating from the proximal nail matrix, which leads to the presence of abundant intracellular vacuoles and to a lesser compactness of keratins. Genes mapping within this chromosomal region include the genes coding for type II (basic) cytokeratins and
P.34
hard keratins, and the gene defect resides on chromosome 12q13 (33).
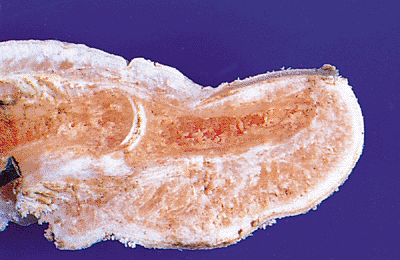 |
Figure 2.6 Sagittal section of an adult thumb, in which it is possible to observe the relations of the nail unit with the adjacent tissues. |
In the proximal portion of the nail, there is an arch called the lunula. The thickness of the nail plate is 0.5 mm in women and 0.6 mm in men (34). The nail plate is delimited by three folds: two lateral and one proximal (Figure 2.9). If the nail plate is avulsed, the grooves become visible where the nail plate rested. These potential spaces are only real spaces in abnormal conditions of the nail, as in paronychia. In the lateral nail grooves, the epidermal lining does not contribute to the formation of the nail plate except in the most proximal portions, where it becomes continuous with the epidermis of the proximal groove or matrix.
The proximal nail fold is the most important one, since, as we shall note later, its contribution to the formation of the nail plate is fundamental (35). This fold shows two portions: a dorsal portion, lodging the matrix, and a ventral portion. Twenty-five percent of the total surface of the nail plate is located under the ventral portion of the proximal nail fold. The terminal tendon of the digital extensor is closely related with this area, and the thin nature and proximity with the nail matrix must be kept in mind during surgery (36).
 |
Figure 2.7 Cross section of an adult finger. The nail plate lies on nail bed, and the lateral border is overlapped by lateral nail fold. |
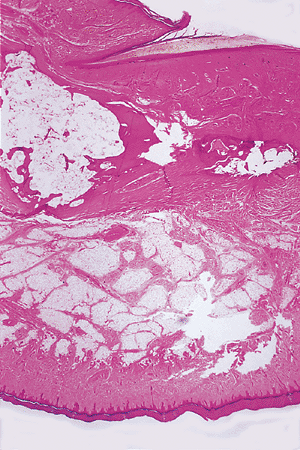 |
Figure 2.8 Histological sagittal section of a finger (H&E, 10). |
A white crescent-shaped lunula can project from under the proximal nail fold. It is usual on the thumbs and common on other fingers and on large toenails. The lunula
P.35
is the most distal portion of the matrix and determines the shape of the free edge of the nail plate. The color of the lunula is partly due to the effect of light scattered by the nucleated cells of the keratogenous zone of the matrix and partly to the thicker layer of epithelial cells making up the matrix (35,37).
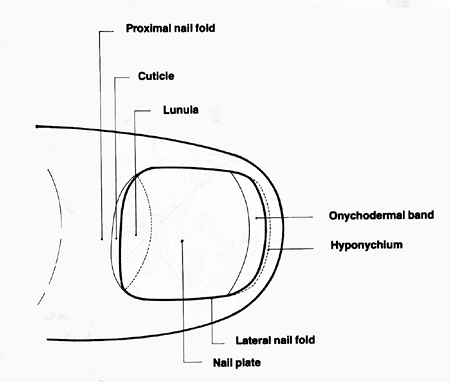 |
Figure 2.9 Schematic nail diagram, including nomenclature. |
At the point of separation of the nail plate and the nail bed, the subungual epidermis may be modified as the solehorn (38). In humans, this structure may only be vestigial, its original significance only being evident from comparative anatomical studies. However, in certain diseases, it could be the seat of distal subungual hyperkeratosis or parakeratosis; for example, in pachyonychia congenita and pityriasis rubra pilaris (39). The distal limit of the ungual layer is the hyponychium, which determines the formation of the distal fold, a keratinized structure that continues until the fingertips. A subungual extension of the hyponichium and obliteration of the distal groove is named the pterygium inversum unguis (40). This term was coined because of the similarity between the behavior of the hyponychium and the eponychium in classic cases of pterygium unguis.
On close examination, two further distal zones can often be identified: the distal yellow-white margin and, immediately proximal to this, the onychodermal band (41). This band is a barely perceptible narrow transverse band, 0.5 to 1.5 mm wide, that is more prominent in acrocyanosis. The exact anatomical basis for the onychodermal band is not known, but it appears to have a different blood supply from the main body of the nail bed (42). It is possible to explore it through a strong compression of the distal zone of the finger, leaving behind a white band. The band's color can occasionally be modified by diseases (39,43).
Several studies have been published about the exploration of the nail apparatus. Although ultrasound transmission can be useful for studying the nail plate thickness (44), magnetic resonance imaging (MRI) permits the detection of subungual lesions smaller than 1 mm in diameter (45).
 |
Figure 2.10 Horizontal section of the dorsal nail plate. Corneocytes show a polydedral disposition with rounded corners; the cells do not contain nuclei or elements (VVG stain, 200). |
Microscopic Anatomy
The Nail Plate
Microscopically, the nail plate consists of closely packed, adherent, interdigitating corneocytes lacking nuclei or organelles (Figures 2.10,2.11). Many intercellular links, including tight, intermediate, and desmosomal junctions, are present (46). The nail plate is made-up of three layers: a thin dorsal layer, a thick intermediate layer, and the ventral
P.36
layer from the nail bed. The cells of the surface of the nail plate overlap, slanting from proximal-dorsal to distal-volar. For this reason, the dorsal surface of the nail plate is smooth, whereas the palmar surface is irregular, showing longitudinal striations. This can also be observed with optical microscopy, scanning electron microscopy (47), and x-ray microdiffraction (48). With this latter technique, Garson et al. (48) demonstrated three different layers in the nail plate that are characterized by different orientations of the keratin molecules from the outer to the inner side of human nail. These layers were associated with the histological dorsal, intermediate, and ventral plates. The hairlike type -keratin filaments (81 in diameter) are only present in the intermediate layer (accounting for approximately two-thirds of the nail width) and are perfectly oriented perpendicularly to the growth axis in the nail plane. Keratin filaments of stratum corneum (epidermis) type, found in the dorsal and ventral cells, are oriented in two privileged directions; parallel and perpendicular to the growth axis. This sandwich structure in the corneocytes and the strong intercellular junctions gives the nail high mechanical rigidity and hardness, both in the curvature direction and in the growth direction. Lipid bilayers (49 thick) parallel to the nail surface fill certain ampullar dilations of the dorsal plate and intercellular spaces in the ventral plate. Using x-ray microdiffraction, Garson et al. also showed that onychomycosis disrupts the keratin structure, probably during the synthesis phase. No keratohyalin granules were seen, but acidophilic masses, called pertinax bodies of Lewis and Montgomery, are occasionally seen in older age groups.
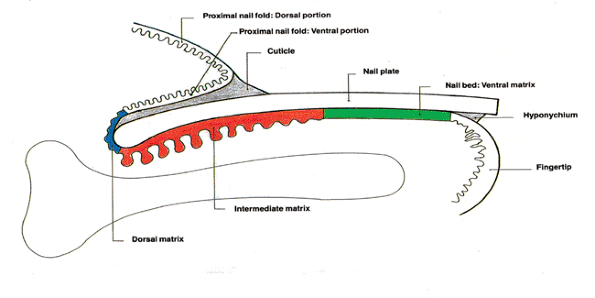 |
Figure 2.11 Schematic diagram of a sagittal section through the nail unit. |
Hamilton et al. (49) believed that the progressive increase in the thickness of the nail plate with age is attributable to the increasing size of the cells in the plate, consecutive to the frictional loss of nail; however, Johnson and Shuster (50) studied 20 normal great toenails to find the determinant of final nail thickness and length at its point of detachment at the onychodermal band. They confirmed that the increase of nail thickness with age is independent of the frictional traumatisms on the plate. Cutting tests showed that fracture of the nail plate occurred because the energy to cut nails transversely, at approximately 3 kJ/m2, was about half that needed (approx. 6 kJ/m2) to cut them longitudinally (51).
Corneocytes of the human nail plate have been studied by Germann et al. (52). Corneocytes of the dorsal nail plates of normal nails are irregular and polyhedral, nonnucleated, and with distinctly irregular networks. These horny cells from nail plates increase in size with age: babies have small cells, adults have significantly larger cells, and aged subjects have significantly larger cells than the younger adults. These authors also commented that the faster-than-normal growing nail plates yield smaller cells; for example, corneocytes from psoriatic patients are smaller than normal, whereas corneocytes from slow-growing nails, such as from persons having lichen planus or dyskeratosis congenita, are larger than normal.
Frequent gap junctions were observed near the area where lamellar granules were discharging their contents, and it was suggested that a certain substance might be able to pass through the nail plate using such intercellular channels. Perhaps such channels explain the greater permeability of the nail plate to polar molecules compared with the permeability of the skin (53). The water content of human nail plates has been determined using a portable near-infrared spectrometer with an InGaAs photodiode array detector and PLS regression by Egawa et al. (54).
Transonychial water loss (TOWL) in vivo has been studied by Nuutinen et al. (55). These authors have demonstrated that TOWL values decrease with age and patients with eczema, psoriasis, and onychomycosis have significantly lowered TOWL values compared with healthy subjects.
The biochemical composition of the nail plate has been widely studied. Calcium, found as the phosphate in hydroxyapatite crystals, is an important component of the nail plate; it is intracelluarly bound to phospholipids, particularly in the dorsal and ventral nail plate (56). Calcium concentration is approximately 0.1% of the weight, 10 times greater than in hair (57). However, some authors believe that the proportion of calcium in the nail contributes little to the hardness of the nail plate in human beings (39,58). Also, it is possible that calcium is not an intrinsic part of the nail but is incorporated from extrinsic sources, such as soaps; the nail is relatively porous, and calcium could enter as ionic calcium or bound to fatty acids. Other metals such as copper, manganese, zinc, and iron are also found in small quantities in the nail plate, although their function is still unknown (58,59). Lipids are also an important component of the nail plate. Helmdach et al. (59) have demonstrated that nail plate lipid composition varies with age and sex: the lipid composition of the fertile years shows distinct profiles compared to that of childhood and old age, suggesting an influence of sex hormones on nail lipogenesis.
The existence of sulfhydryl and disulfide groups has been demonstrated in the nail plate. During early embryonic life, there is a very high concentration of the sulfhydryl groups (9), which decreases as the delivery date approaches and stabilizes at about the age of 3 years (14). These sulfurous radicals are formed at the expense of amino acids, such as cystine. Quantification of cysteine and cystine can be performed by hydrolysis (60). Total sulfur concentration is similar in the dorsal and intermediate plates. Compared with hair, the nail plate also contains glutamic acid, serine, and less tyrosine (57,58,59,60).
In certain diseases, the quantity of various organic and metallic components of the nail plate can be increased. A brief listing is presented for reference: total nonprotein nitrogen, urea nitrogen, ammonia nitrogen, and uric acid in gout (61); creatinine in chronic renal failure (62); sodium in cystic fibrosis (63,64); calcium in older subjects (48,57);
P.37
copper in Wilson's disease (65); arsenic as biomarker to arsenic exposure in the endemic areas (66); and morphine, 6-acetylmorphine, and cocaine in drug abusers (67).
An analysis of the keratin of the nail plate revealed the following (68): (a) -fibrillar, low-sulfur protein; (b) globular, high-sulfur matrix protein; and (c) high-glycine tyrosine-rich matrix protein. All these fractions are also present in hair. The hardness of nail is due to the high-sulfur matrix protein, contrasting with the relatively soft keratin of the epidermis.
Proximal Nail Fold
The proximal nail fold is an invaginating, wedge-shaped fold of the skin on the dorsum of the distal digit, and the nail plate arises from under this fold (Figure 2.12). The proximal nail fold consists of two layers of epidermis: the ventral portion overlying the newly formed nail plate and the dorsal portion that forms the dorsum of the finger epidermis. The keratinization process in both portions does not differ from that of the epidermis elsewhere, possessing a granular layer that is absent in all parts of the nail matrix.
The dorsal portion of the proximal nail fold consists of a continuation of the epidermis and dermis of the dorsal digit with sweat glands but no follicles or sebaceous glands. At the distal tip, a thick corneal layer called the cuticle shows on the dorsal surface of the nail plate (Figure 2.13). Its function is the protection of the nail base, particularly the germinative matrix. Loss of the cuticle often allows acute and chronic inflammatory and infective processes to involve the nail matrix, leading to secondary nail plate dystrophies.
 |
Figure 2.12 Proximal nail fold with its two portions: dorsal portion, with identical histologic pattern to the skin of the dorsum of the distal digit; ventral portion, overlying the nail plate. Note the great thickness of the stratum corneum of this epithelium (MF stain, 63). |
 |
Figure 2.13 Detail of the cuticle. At the distal tip, the proximal nail fold shows a thick corneal layer (cuticle) on the dorsal surface of the nail plate. |
The ventral portion is thick-skinned, has no appendages, and is closely attached on the dorsal surface of the nail plate. The epithelium of the ventral surface of the proximal nail fold has been called the eponychium (9,37,39). Diseases that affect the ventral portion of the proximal nail fold can affect the newly formed nail plate. For this reason, some authors think that the proximal nail fold contributes to form the superficial layer of the nail plate. In particular, the apparition of pits and grooves (Beau's line) on the nail is due to parakeratotic and growth detention phenomena, respectively, in the ventral portion of the proximal nail fold.
Matrix
The ventral surface of the proximal nail fold forms the roof of the proximal nail groove; the nail matrix forms its floor, and the nail plate lies between the two. The matrix is divided into three parts (8,9,39): dorsal, intermediate, and ventral. Of these, the dorsal and, above all, the intermediate portions play an important role in nail plate formation. In particular, the true matrix is the intermediate portion. For this reason, when we discuss the histology of the matrix, we are fundamentally referring to the intermediate portion. The ventral portion corresponds to the nail bed; the controversy about its participation in the formation of the definitive nail plate is discussed in the section entitled Nail Growth.
The main body of the matrix is composed of epithelial cells, with melanocytes, Merkel cells, and Langerhans cells scattered among the epithelial cells.
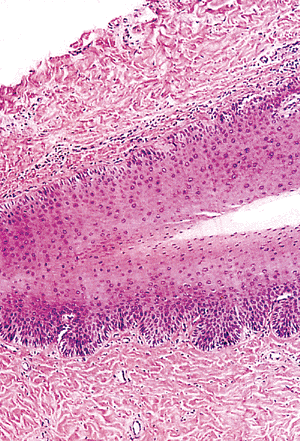 |
Figure 2.14 Histologic appearance of the matrix angle, formed by the ventral portion of the nail fold and the dorsal and intermediate matrix. |
P.38
Epithelial Cells
The matrix is an easily identified thick squamous epithelium, situated immediately below the ventral portion of the proximal nail fold (Figure 2.14). Its main feature is its thickness, with between 8 and 15 mamelons (protuberances) (Figure 2.15). Its undulation can be seen for only a few millimeters, flattening itself in the area corresponding to the nail bed. As in the epidermis of the skin, the matrix possesses a very active germinative basal layer of immature basaloid cells, producing keratinocytes which differentiate, harden, die, and contribute to the nail plate (Figure 2.16). The nail plate is formed by a process that involves flattening of the basal cells of the matrix, fragmentation of nuclei, and condensation of the cell cytoplasm to form horny flat cells. An important histologic feature is the lack of granular layer. Acanthosis and papillomatosis are only seen in the nail unit in the matrix and, distally, in the hyponychium (Table 2.2).
Table 2.2 Histologic Characteristic Features of each Zone of the Nail Unit | ||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
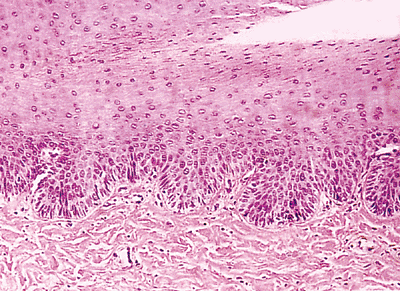 |
Figure 2.15 Detail of matrix epithelium. This zone shows an acanthotic epithelium, with germinative basal cells keratinocytes and scarce melanocytes. |
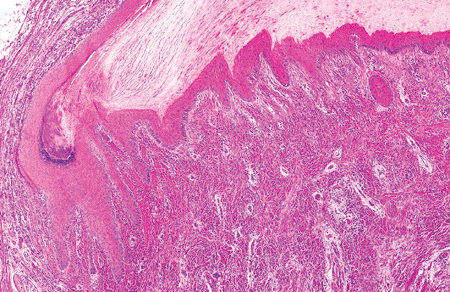 |
Figure 2.16 Detail of the matrix zone, in which one can observe the abrupt keratinization. |
P.39
Melanocytes
The nail matrix possesses melanocytes, just as the hair matrix does. The matrix of Caucasian patients contains sparse, poorly developed melanocytes (Figures 2.17, 2.18, 2.19) (69). It is difficult to observe melanocytes in the proximal matrix zone using light microscopy, but their number is progressively increased distally. Nevertheless, the number of melanocytes is always less than in normal skin (69,70,71,72,73).
There are distinct differences in the distribution of melanocytes in adult skin and nail matrix. Immunostaining of nail matrix melanocytes revealed that they are not singly interspersed between the keratinocytes of the basal layer, but they are frequently arranged in small clusters among the suprabasal layers of the nail matrix (73,74). A similar pattern of distribution of melanocytes has been described in fetal skin and in fetal skin equivalents, in which the melanocytes are grouped and localized both basally and suprabasally. The suprabasal location of nail matrix melanocytes may be a consequence of differences in the distribution of adhesion molecules in the nail epithelium (73,74).
Higashi and Saito (75) demonstrated that the number of melanocytes and the intensity of the dopa reaction in them were much greater in the distal than the proximal matrix. The melanocyte count in normal epidermis was reported to be 400 per 2.784 mm2 (74,75), while the range was 208 to 576 in the distal areas of the intermediate nail matrix (75).
Ultraviolet rays and trauma are factors that could influence a more extensive distribution in the distal zone (73,74). In some races such as Japanese, the matrix contains several hundred well-developed melanocytes per millimeter (75). Also, it seems that melanocytes of the nail matrix in Oriental races have larger dendritic processes than Caucasians.
 |
Figure 2.17 Observe the notable hyperpigmentation of the basal layer in the pulp of the finger in contrast with the absence of pigmentation in the nail matrix (Fontana's stain, 4). |
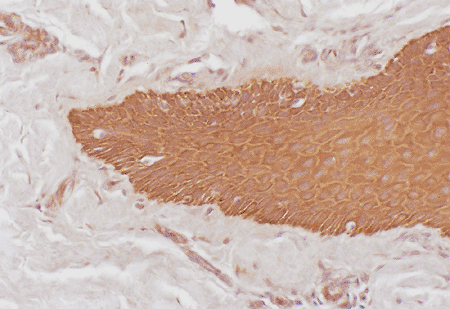 |
Figure 2.18 Melanocytes of the nail matrix, are negative with cytokeratin antibody stain. Observe its scarce number in the nail matrix (cytokeratin antibody, 10). |
Pigment, therefore, arrives in the nail plate as in the keratinized cells of the stratum corneum and hair cortex (76). Nail pigmentation is most evident in African Americans, in whom it is commonly seen as longitudinal linear streaks, although this anomalous distribution of pigmentation can also be seen in pathological states such as subungual pigmented nevi and melanomas in the matrix zone (77). Abundant melanosomas of these subjects have a protective effect against ultraviolet radiation, since variations in racial pigmentation are due to the number and size of melanosomes produced (78).
The location of melanocytes in the matrix is directly related to the location of pigmented bands, most of which originate in the distal matrix and do not cross the lunula (73). Longitudinal melanonychia in AIDS patients is a common finding. Although this pigmentary nail change
P.40
can be related to the zidovudine oral treatment, in other cases it seems to be independent and associated with elevated levels of -melanocyte-stimulating hormone ( -MSH) (79). Cancer chemotherapeutic agents often induce nail changes, and nail pigmentation is probably the most frequently observed nail abnormality. The mechanism of drug-induced hyperpigmentation is not fully understood, but this is most probably due to increased melanogenesis in matrix melanocytes. Beau's lines, transverse leuconychia, and onychomedesis are the result of nail matrix toxicity and can be seen after intensive or combined chemotherapy. Nail bed toxicity, as manifested in apparent leuconychia or onycholysis, may be seen in association with nail bed hemorrhage, which may be exacerbated by concurrent thrombocytopenia. Acute paronychia has also been described after treatment with methotrexate (80).
 |
Figure 2.19 Melanocytes of the lunula, not staining with cytokeratin antibody in contrast with positive epithelial cells. Note its location in basal and suprabasal epithelial layers (cytokeratin antibody, 20). |
Langerhans and Merkel Cells
Langerhans and Merkel cells have also been identified in the matrix (70), although their signification is unknown. Studies of the Langerhans cells in the nail matrix are almost absent. Nevertheless, interesting studies about the Merkel cells have been recently published. Moll and Moll (81) studied the Merkel cells (MCs) in ontogenesis of human nails, using immunohistochemical stains with cytokeratins 18 and 20 in human fetuses of 9 to 22 weeks of life. These authors have concluded the number of MCs are detected very early (nine weeks) in the matrix primordium. However, MCs were found to decrease in number with aging of the fetuses: MCs were only seen in the proximal nail fold at 12 to 15 weeks and were essentially absent from the epithelium of the ventral matrix and nail bed in the adult.
Lunula
The intermediate matrix continues forward with a visible, white half-moon-shaped area called the lunula. The lunula is shown to be linked to a well-defined area in the underlying dermis with a specific histology and microvascularization. Although always present, it cannot be seen in some fingers but is most visible in the thumbs. The typical white color is related to some histologic features of this area. Lewin (34) confirmed that the opacity of the proximal nail plate, the relative avascularity of the subepidermal layer, and the loose texture of the dermal collagen are responsable for its color. Samman (82,83) thought that it was a combination of incomplete keratinization in the nail plate and loose connective tissue in the underlying tissue. Zaias (84) believed that the nail plate would be thinner in the lunula because it coincides with the keratogenous zone, the zone of cytoplasmic condensation in the matrix just before cells form the nail plate. The length of the subnail matrix area distal to the free edge of the proximal nail fold is highly correlated with the length of the lunula (85).
Other special histologic features of this zone of the matrix, including a different chemical composition of the nail plate and a different distribution of the dermal fibers, have been related to the typical white color of the lunula (86,87), although not one of these factors has been confirmed. We do not even know the exact function of the lunula.
Nail Bed
The nail bed begins where the intermediate matrix ends, and some authors prefer to designate the ventral matrix as the site (39,86). A histologic appreciation of the end of the intermediate matrix and the beginning of the nail bed is very easy. The nail bed epidermal layer is usually a flat epithelium no more than three- or four-cells thick and without melanocytes (Figures 2.20,2.21,2.22). The transition zone from living keratinocytes to dead ventral nail plate cells is abrupt, occurring in the space of one horizontal cell layer in a manner very similar to what occurs in the Henle layer of the internal root sheath of the hair follicle (87).
During its early development, the nail bed exhibits a keratinization process differing from the adult's, with a prominent granular layer at 17 to 20 weeks of development. However, after birth, the nail bed, like the matrix, keratinizes without a granular layer. lt is less active than the matrix, with a longer turnover time than the matrix and skin (88). A thin parakeratotic keratin is produced, apparently dragged forward by the nail plate growing over it rather than becoming incorporated into the nail.
In the nail bed, the dermis fits into the longitudinal and parallel nail bed ridges in tongue-and-groove fashion. The fine capillaries of the nail bed run in these parallel dermal ridges, and disruption of these accounts for the splinter hemorrhages commonly seen in normal and disease states (42). There is no fat tissue in the nail bed, although scattered dermal fat cells may be visible microscopically.
 |
Figure 2.20 Nail bed. Note the flat epithelium with an interdigitated upper zone. |
P.41
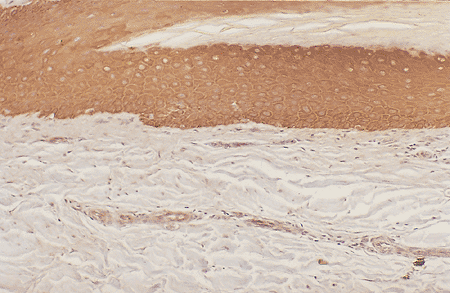 |
Figure 2.21 Melanocytes are absent in the nail bed (cytokeratin antibody stain, 10). |
The nail bed epidermis moves distally toward the hyponychium. The cells that appear to be the germinative population lie near the lunula, so close together that they may be confused as belonging to one population. The distal movement from this position may also help explain why during development the nail bed epidermis seems to lose keratohyalin granule layers from a proximal-to-distal direction concomitantly with the formation of the primitive nail plate (89).
In some pathologic states, the nail bed shows a granular layer in which the activity in the nail bed is greatly increased, such as occurs in onychogryphosis, pachyonychia congenita, and psoriasis (90); in these cases, the horny cells produced push the nail plate upward and give it a claw-like appearance.
Table 2.3 Histochemistry of the Naila | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
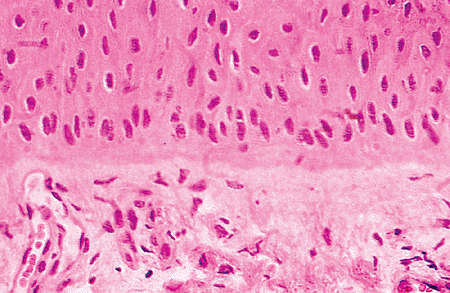 |
Figure 2.22 Detail of the nail bed zone. The epithelium shows a few active germinative cells at the basal layer. In the upper dermis, it is possible to observe larger vessels than in the normal skin. |
Histochemical studies of the nail bed prove the presence of bound phospholipids in the nail bed epidermis (Table 2.3). Bound cysteine can be detected in the transition zones: acid phosphatase and nonspecific esterase are absent in the dorsal and intermediate zones (39,56).
Immunohistochemical studies have demonstrated that nail bed expressed all the target antigens found in the normal nonappendageal basement membrane. In particular, there was normal expression of the epidermal-associated antigens, the 230- and 180-kDa bullous pemphigoid antigens, and the 6 4 integrin. There was also normal expression of the lamina lucida antigens LH39, GB3, and laminin. Sinclair et al. (91) pointed out that the dermal-associated components, namely the 285-kDa linear immunoglobulin A (IgA) antigen, the extracellular matrix glycoproteins chondroitin sulphate, type VII collagen and its closely associated proteins, and the poorly characterized antigen for LH24 and LH39 were all normally expressed. All the former molecules were also found in the proximal nail fold, nail matrix, and hyponychium.
The presence of antimicrobial peptides in nails (mainly cathelicidin LL-37, demonstrated by immunostaining and which has activity against relevant nail pathogens) may account for the ability of the nail unit to resist infection
P.42
in the absence of direct access to the cellular immune system (92).
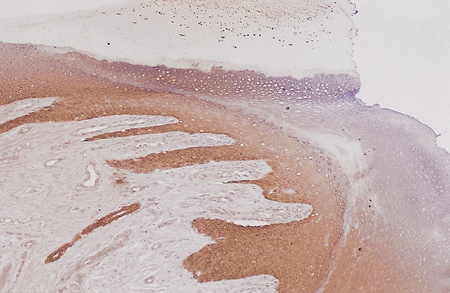 |
Figure 2.23 Panoramic view of the hyponychium (cytokeratin stain, 10). |
Hyponychium
The most distal portion of the nail bed is the hyponychium, representing the union between the nail bed and the fingertips (Figure 2.23); its histologic characteristics are rather peculiar. This transition zone presents a notable change of appearance after a few millimeters because the epithelium undergoes keratinization similar to that of the epidermis (Figure 2.24). The result is marked acanthosis and hyperplasia with the crests oriented almost horizontally; this is associated with normal appendages (Figure 2.25). An area of abundant keratohyalin granules is present, and the horny layer produced tends to accumulate under the free edge of the nail plate, producing a keratin horn similar to the cuticle. The hyponychium is the first site of keratinization in the nail unit (8,9,11,12,13) and of all epidermis in the embryo (93). The function of this anatomical formation is to render the nail bed impermeable to protect it from external agents (94). If this structure fails, dermatophyte invasions will be frequent, producing onychomycosis (95).
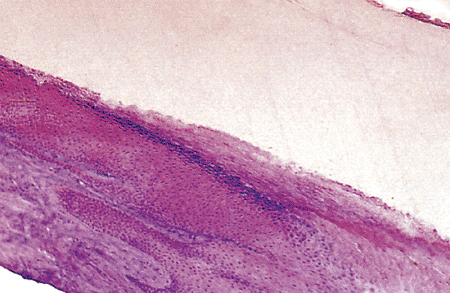 |
Figure 2.24 Hyponychium zone. The most important feature of this zone is the great accumulation of keratin under the distal nail plate. |
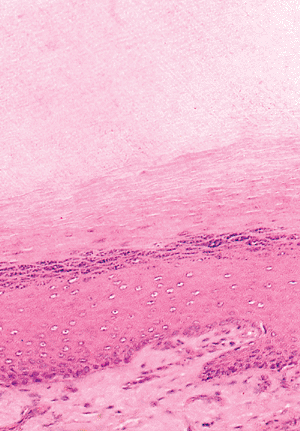 |
Figure 2.25 Detail of the hyponychium zone. Note the great keratin layer under the nail plate and the visible granular layer. The epithelium shows an acanthotic aspect, with transversal papillae. |
Terry (41) describes an intermediate zone between the nail bed and the hyponychium, which he called the onychodermal band. Terry speculated that this area, normally from 0.5 to 1.5 mm wide, had a blood supply different from the remainder of the nail bed, a fact later confirmed by other authors (42). For this reason, the color is paler than the pink nail bed and has a slightly amber tinge with a translucent quality. The onychodermal band occasionally changes its color, especially in cirrhosis and other chronic diseases (39,43).
Lateral Nail Folds
The lateral nail folds have a structure similar to the adjacent skin but are normally devoid of dermatoglyphic markings and folliculo sebaceous units. Acanthosis and hyperplasia of the epithelium are present, similar to that of the hyponychium. Keratinization within the nail folds proceeds by keratohyalin formation in the granular layer (Figure 2.26). The epidermal lining of these grooves does not contribute to the formation of the nail plate except in the most proximal portions of the grooves, where it becomes continuous with the epidermis of the proximal nail fold or matrix.
When the lateral border of the nail plate pathologically breaks this fold, abundant granulation tissue forms, constituting the onychocryptosis, a frequent pathologic alteration of the great toenail.
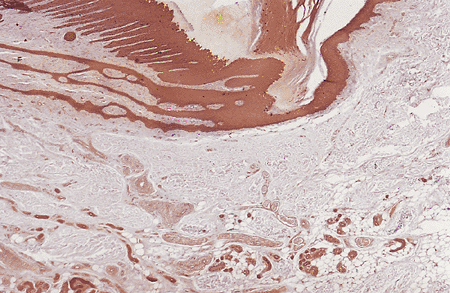 |
Figure 2.26 Lateral nail fold. Observe its acanthotic epithelial layer and the presence of eccrine glands in the middle dermis (cytokeratin antibody, 4). |
P.43
Immunohistochemistry of the Nail Unit
Nail Plate
The cornified envelope of the epidermis is formed by several precursor proteins, including involucrin, keratolinin, loricrin, pancornulin, sciellin, 195-kDa protein, keratin, and filiagrin. Baden and Kvedar (96) used monoclonal antibodies to demonstrate that pancornulin is present in the nail fold and proximal matrix, and sciellin is detectable in the nail fold, matrix, and bed. Similarly, in studies of human nails (which contain hard keratins), the use of immunofluorescence, immunoblotting, and PCR (polymerase chain reaction) have shown that trichohyalin, a 200-kDa protein of the inner root sheath and medulla, was present in the ventral matrix but not in the nail bed; a few scattered cells stained for trichohyalin were observed within the nail plate (97).
Heid et al. (98) studied the keratin expression patterns observed in the human fetal nail matrix and revealed that the nail develops from both skin- and hair-type differentiating cells. Kitahara and Ogawa (99) demonstrated that AE13 antibody reacted with the dorsal nail matrix. Because AE13 antibody recognized hard keratins that are characteristic of differentiation in hair, these results show that adult nail develops in such a way that hair-type differentiation is confined to the ventral nail matrix, supporting the Heid et al. study's results (100,101).
Keratinocytes
Expression of keratins in the different compartments within the nail unit have been demonstrated in some recent articles (102,103,104,105,106,107,108,109,110,111,112). The characteristics of the different keratins found at different sites could be relevant to our understanding of the biology of the normal nail and changes seen in several diseases.
Analysis of human nail plate by gel electrophoresis demonstrates a range of keratins of two characteristic types, as we commented formerly (see Genetics and Nail Keratins). Soft, or epithelial, keratins represent the major structural intermediate filament isolated from human skin but constitute only 10 to 20% of the keratin found in nail (99). Hard keratins, characteristic of hair and nail differentiation, exist in the same acid-base heterodimer configuration as soft epithelial keratins but have additional resistance. Molecular classification of hard keratin proteins on gel electrophoresis describes eight major (Ha1 4 and Hb1 4) and two minor (Hax and Hbx) proteins, all of which are probably present in human nail (103). This family has been extended by genomic analysis into at least seven type I Ha keratins and six type II Hb keratins. Further keratins and their isoforms are likely to be discovered. In addition to the hard keratins, epithelial keratins isolated from the nail plate include K1, K10, K5, K14, K6, K16, K17and K19 from fetal nail (102,103) (Figure 2.27).
Ha1 is one of the major hard keratins found in nail, where hard keratin represents 80 to 90% of nail keratin. Keratins K5, K14, K1, and K10 were also detected in the nail. Since Berker et al. (102) have found a low expression of the differentiation-specific keratins K1 and K10 in the keratogenous zone of the ventral matrix, this feature was not demonstrated by other authors (104).
Keratins K6, K16, and K17 are normally found in hyperproliferative epidermis, such as in psoriasis or in wound healing (105). Studies of proliferative compartments in the nail unit suggest that the nail bed is not a major contributor to the nail plate. It may be that the ventral aspect of the proximal nail fold and not the nail bed is the source of nail plate K6 and K16, and the matrix and not the nail bed provides K17. Keratin K14 is synthesized in the basal layer, and K14 protein was detected throughout the epithelium, as has been noted in other tissues. However, the marker of basal keratin conformation, LH6, was also seen throughout the nail bed. This is unusual and may reflect the absence of the expression of the suprabasal keratins K1 and K10, which are thought to obscure the epitopes detected by LH6 in normal stratified epithelium. This persistence of LH6 antigen is also seen in the outer root sheath of the hair follicle; because this is also the site of expression of K16, K6, and K17 (106), the data support the analogy drawn between the nail bed and outer root sheath (107). However, expression of K1 and K10 is found to a degree in the upper outer root sheath superficial to the level of the sebaceous gland.
The absence of K1 and K10 from the nail bed correlates with a reduction in terminal differentiation. Lack of cornification is also seen in mucosal epithelium in combination with the presence of K16 and K17. However, mucosal differentiation is defined by the presence of K4 and K13, which was absent in the nail bed (108). The exact significance of these keratins remains to be established.
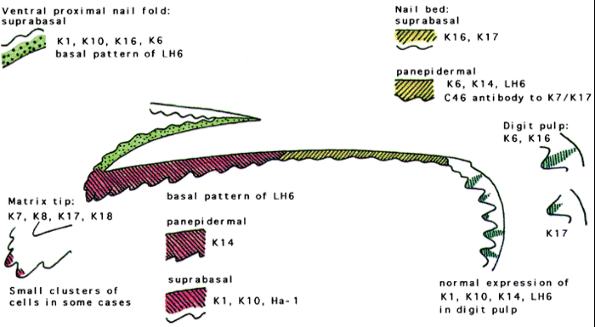 |
Figure 2.27 Keratin expression in the normal nail unit: markers of regional differentiation. Taken from De Berker D, Wojnarowska F, Sviland L, Westgate GE, Dawber RPR, Leigh IM. Keratin expression in the normal nail unit: markers of regional differentiation. Br J Dermatol 2000;142:89 96. |
P.44
Additional molecular studies have demonstrated the homology between hair and nail keratins (109). Retinoic acid inducible gene-1 was originally identified as an orphan G-protein coupled receptor (oGPCR) induced by retinoic acid. Three highly homologous oGPCR (GPCR5B, GPCR5C, and GPCR5D) have since been classified into the RAIG1 family. Inoue et al. (109) studied the unique tissue distribution of GPCR5D and its mechanism of expression by in situ hybridization in hair shafts, nail, and filiform papillae of the tongue. They found that GPCR5D is expressed in differentiating cells that produce hard keratin, including cortical cells of the hair shaft, the keratogenous zone of the nail, and in a central region of the filiform papillae of the tongue. GPCR5D transcript is expressed in hair follicles during mid- and late anagen, and catagen but not at telogen and early anagen phases. The differentiation-inducer, all-trans retinoic acid, induces GPCR5D expression in cultured hair bulb cells. Because the tissue distribution of GPCR5D indicates a relationship with hard keratins that constitute the major structural proteins of hard epithelial tissues, they investigated the effect of GPCR5D on acid hard keratins.
Analyses of cultured cells showed that transient overexpression resulted in suppression of Ha3 and stimulation of Ha4 hair keratin gene expression. The expression was maintained in the hair follicles of whn-deficient (nude) mice, suggesting that this gene is regulated by a signal pathway different from that of hair keratin synthesis. These data provide a framework for understanding the molecular mechanisms of GPCR5D function in hard keratinization.
For the expression of hard keratin by matrical keratinocytes is primordial the influence of matrical fibroblasts. Okazaki et al. (110) demonstrated that, even in non nail-matrical keratinocytes, expression of hard keratin could be induced by nail-matrical fibroblasts. These investigators constructed three different skin equivalents: (a) ventral keratinocytes (from the ventral side of the digit) were cocultured with ventral fibroblasts (group A); (b) ventral keratinocytes were cocultured with nail-matrical fibroblasts (group B); and (c) nail-matrical keratinocytes were cocultured with ventral fibroblasts (group C). Immunohistochemical examinations with antihard keratin antibody (HKN-7) revealed hard keratin expression in groups B and C. The HKN-7-positive cells were distributed continuously in the entire epithelial strata or in the suprabasal layer in group B, whereas HKN-7-positive cells were distributed spottily in group C. This study indicated extrinsic hard keratin induction in non nail-matrical keratinocytes by nail-matrical fibroblasts and suggests that non nail-matrical epidermal grafts may be effective in the treatment of de-epithelialized nail injuries. In addition, it is possible that lost nails could be reconstructed with grafts of tissue-engineered nail equivalent.
Human carcinoembryonic antigen (CEA) and CEA-related molecules play an important role in adhesion of the nail plate to the nail bed. Egawa et al. (111) demonstrated that a CEA family antigen with NCA (CD66c)-like immunoreactivity was strongly expressed in the nail. A strong expression of the NCA-like antigen was only seen in the keratinocytes distributed in the upper epithelial cell layers of the major central portions of the nail bed, to which the nail plate is firmly bonded; expression was stronger at the more distal portion of the nail bed and was absent in the nail matrix, the hyponychium, and the lateral folds. The results are interesting because the nail plate is firmly bonded to the nail bed, less so proximal to the matrix margin, and it has been shown that the bed epithelium travels at the same speed as the nail plate, indicating that the bed epithelium has a proximal site of origin and a distal end.
Plasminogen activator inhibitor type 2 (PAI-2) was detected in the differentiating cells of the matrix and nail bed (112). These authors, using transfected cell lines that express high levels of PAI-2, have suggested that this inhibitor may confer protection against programmed cell death. This consistent, selective distribution of PAI-2 in the postmitotic, maturing cells prior to terminal keratinization and death suggests that (a) PAI-2 may be considered as a differentiation marker for many epithelial cell types, and
P.45
(b) PAI-2 is appropriately positioned to protect epithelial cells from premature demise (112).
Merkel Cells
Lacour et al. (113), in a double indirect immunofluorescence and immunoelectron microscopic study with the monoclonal antibody Troma-1, have only found MCs in the proximal nail fold of the adults, with a concentration greater than 50 MCs/mm2.
Immunohistochemically, keratins K8 and K18 have been used as markers of MCs (81). Merkel cells have neuroendocrine characteristics and are of uncertain function, although their prominence in the nail unit in the early stages of fetal development has been noted, and a role in ontogenesis has been proposed. The number and location of cells demonstrating K8 and K18, which included the rete ridges of the digit pulp, suggested that these cells were MCs rather than they contributing directly to nail plate formation.
Melanocytes
Tosti et al. (114) have recently studied the melanocyte characterization of the normal nail matrix using immunohistochemistry techniques. These authors found nail matrix melanocytes reacted with the antibodies anti-PEP-1, anti-PEP-8, and anti-TMP-1, which recognize the tyrosinase-related protein-1, the tyrosinase-related protein-2 (DOPA-chrome tautomerase), and the tyrosinase-related protein encoded by pMT4 (Table 2.4). This confirms that, even if normally quiescent, nail matrix melanocytes possess the key enzymes responsible for the formation of melanin pigment (115).
Expression of integrins in the nail matrix have been studied by Cameli et al. (116). These authors found that 2 1 and 3 1 expression differ in nail matrix epithelium. In the nail matrix, these integrins are not only expressed on the basal layer, but also on the fourth to fifth suprabasal layers, with suprabasal expression gradually decreasing from the distal to proximal matrix (Table 2.5). As in the normal human epidermis, 1, 4, and 5 integrins subunits are not expressed in the nail matrix; in the same way, ICAM-1, the ligand of LFA1, was negative in the matrix cells. The expression of 1 subunits in the suprabasal layers of the nail matrix indicates a very strong cohesion between nail matrix cells, thus probably revealing them to be an essential prerequisite for the development of a compact nail plate. Cultures of nail matrix cells may represent a useful model to study the biologic properties of nail structure (117).
Table 2.4 Immunostaining of Human Nail Melanocytesa | ||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||
Ultrastructural Anatomy
Very few studies of the normal ultrastructural morphology of the nail exist (10,11,12,13,46,47,69,70,71,72,76,89) because of varied difficulties (70): (a) achieving proper fixation and
P.46
adequate penetration of epoxy resin into the nail plate; (b) obtaining ultrathin sections; and (c) securing the high-voltage electron beam necessary to penetrate through extraordinarily hard tissue (because of the availability of 100 to 200 kV machines).
Table 2.5 Integrin Expression in Human Nail Matrixa | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
The proximal end of the human toenail is composed of several layers of epithelial cells. Hashimoto and coworkers (69,70,71,72) make the distinction between a proximal dorsal, apical, and ventral matrix, although noting that there are few differences between them. They found that the cells composing the proximal matrix were: (a) relatively small, elongated basal cells attached to the basal lamina; (b) relatively large, round, or polygonal squamous cells filling the more central portion of the matrix; (c) melanocytes; (d) Langerhans cells; and (e) Merkel cells.
Moreover, there exists a system of attachment to the dermis, showing the surface of the basal cell with frequent fingerlike elongations that interdigitate with the papillary dermis (Figure 2.28). This results in the formation of numerous micropapillae, with bundles of very fine fibrils (11 to 12 m). The subjacent dermis of the matrix zone shows poor vascularity and scarce collagen fibers, with abundant basic matrix.
The basal cells are very active, with frequent mitotic figures. They showed an elongated nucleus and cytoplasm with numerous, slender projections (villi) intricately interdigitated with neighboring cells. Tonofibrils were also seen as a perinuclear ring with an interposition of the nuclear clear zone in which the majority of mitochondria, transferred melanosomes, and occasional centrioles are located. The suprabasal matrix cells are also round, with frequent mitotic figures. Generally, the long axes of these cells were oriented axiodistally, suggesting the direction of their migration. Large intercellular spaces were often seen between these suprabasal cells. The extensive interdigitation of the peripheral villi as seen in the basal cells disappeared, and multiple desmosomal junctions alone connected these cells (Figures 2.29 and 2.30).
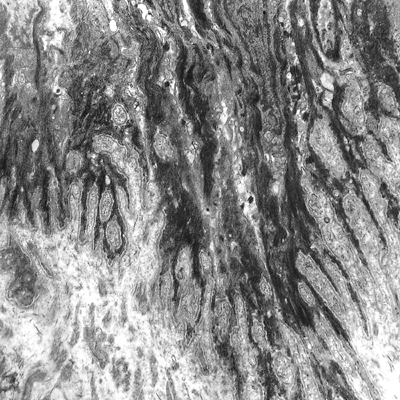 |
Figure 2.28 Ultrastructural appearance of the dermoepidermal junction of the intermediate matrix. The basal layer shows an accentuated digitiform distribution, with multiple intermediate filaments ( 7000). |
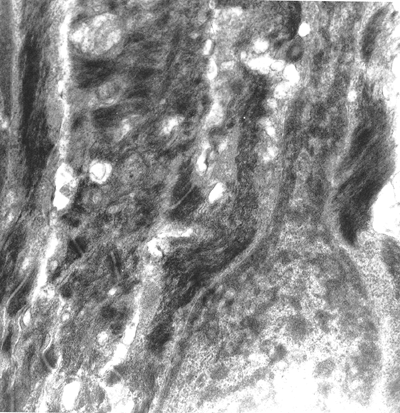 |
Figure 2.29 Detail of the basal layer of the matrix. Extensive condensation of cytokeratin. The nuclei show the habitual shape of normal skin at this epidermal level ( 12,000). |
Abundant desmosomes can be seen in the intermediate layers with high condensations of intermediate fibrils. The aspect of the intermediate layer of the nail matrix is similar to the upper layers of the normal epidermis (Figure 2.31). The cells have lost their organelles, and their cytoplasm is nearly filled with tonofibrils. For this reason, the keratinization process is very abrupt, passing from three to four cellular lines to completely keratinized corneocytes.
Other Tissues of the Nail Unit
Dermis
The dermal component of the nail structures is a very specialized tissue, unique in that it is limited by the underlying phalanx and closely associated with its vasculature and nerve supply. There is no subcutaneous tissue, as previously noted.
Dermis, epithelium, and nail plate in the nail bed present special histologic features due to the great traction that is supported. The dermis is very thick with a dense collagen layer. These fibers are vertically situated in the proximal zone of the nail bed (Figure 2.32) and inclined at an angle of 45 degrees in the zone adjacent to the hyponychium (Figure 2.33).
P.47
Their mission is to attach the nail plate directly with the phalangeal periosteum. Conversely, the nail plate and nail bed are quite firmly attached to each other, more so than the nail plate to the matrix, and this seems to be accomplished by the striking, deep longitudinal ridges and furrows of the nail surface of the nail plate. The nail bed has a unique, longitudinal, tongue-and-groove spatial arrangement of papillary dermal papillae and epidermal rete ridges. This feature is easily observed in transverse sections, in which this arrangement is appreciated as a serrated interdigitation of the ventral surface of the nail, papillae, and rete. These furrows can be seen very well macroscopically just after avulsion of the nail plate, but they are also beautifully shown microscopically by a scanning electron microscope (Figures 2.34 and 2.35) (89).
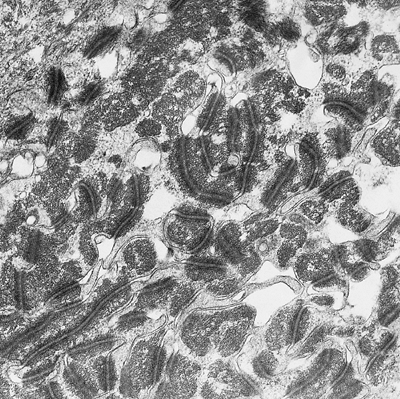 |
Figure 2.30 Detail of the desmosomal junctions of the suprabasal layer. They are bigger and more abundant than in a normal epidermis ( 12,000). |
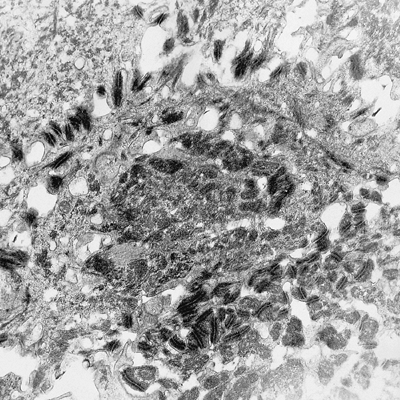 |
Figure 2.31 Intermediate stratum of the matrix epithelium. The intermediate filaments of cytokeratin show a special disposition in the nail matrix ( 4400). |
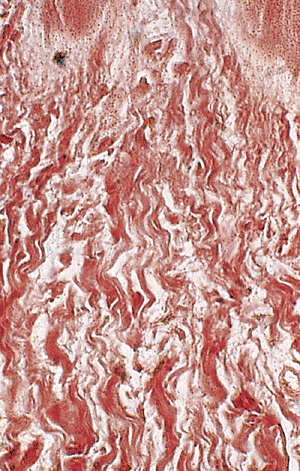 |
Figure 2.32 Collagen fibers of the proximal nail bed. Observe the peculiar vertical disposition (reticulin stain, 400). |
There are few studies about the nerve supply of the nail. The matrix and nail bed present sparse nerve endings and few Vater-Pacini (118) and Meissner corpuscles (119). Intraepithelial nerve fibers were described at the beginning
P.48
of this century (118), but other authors (120) were unable to confirm the description. Experimental studies have established that, for digit-tip regrowth, the major nerve supply is not needed but the nail organ is needed. Nail organs of regrowing fetal digit-tips have been shown to produce Msx, a transcription factor associated with limb bud outgrowth (121). In the same way, it is plausible that nail bed epithelium, acting like an inductive wound epithelium in newts, provides growth factors in mammals that can substitute for those generated by nerve in newt limbs (124).
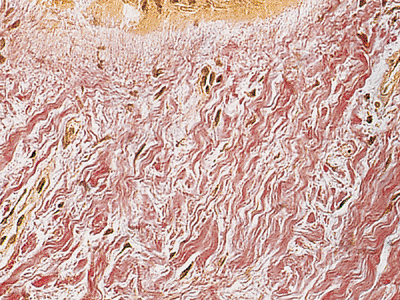 |
Figure 2.33 Collagen fibers of the distal nail bed. Observe the peculiar inclined disposition (VVG stain, 200). |
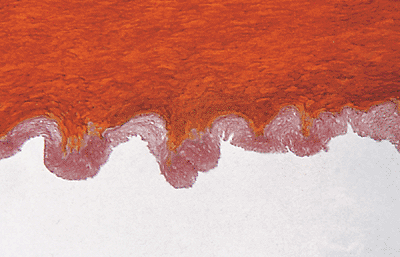 |
Figure 2.34 Transverse section of the nail plate. Note the serrated lower surface of the nail plate (MF stain, 200). |
The hyponychium is the area with greater abundance of nerve endings of the nail and with abundant Meissner and Merkel-Ranvier corpuscles, as in the lateral nail folds (120). This histologic feature gives the hyponychium an important role in the fine sensibility of the finger.
Bone
The nail apparatus includes the subjacent bone. Although the bone has been ignored during the last years, a recent study of postamputational repair following digit-tip amputation revealed an unexpected correlation between nail regrowth and bone regrowth. In this way, Zhao and Neufeld (123) have studied this relationship, observing that in the absence of nail, bone did not regrow at distal levels, and, conversely, when the nail was surgically retained, bone regrew from proximal levels.
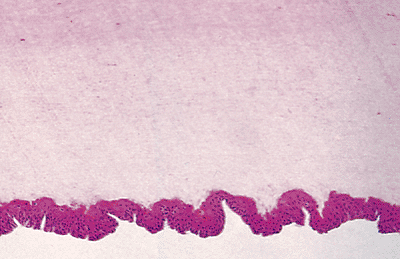 |
Figure 2.35 Avulsed nail plate. Observe the sinusoidal form of the nail bed epithelium attached to the lower surface of the nail plate. |
Blood Supply
The nail has a rich vascularization that deserves separate mention. The arterial blood supply of the nail bed and matrix is derived from paired digital arteries. The most important studies have been published by Flint in 1955 (124), Ryan in 1973 (125), and Smith et al. in 1991 (126), concluding that the main supply passes into the pulp space of the distal phalanx before reaching the dorsum of the digit. An accessory supply arises further back on the digit and does not enter the pulp space. The digital arterial system manifests three characteristic anatomic features: arched anastomotic arteries in the deep dermis; more superficial terminal arteries branching to supply the rete (127), and the great tortuosity of the arterial architecture subjacent to the nail apparatus (Figure 2.36). The arteries possess inner longitudinal and outer circular coats of smooth muscle (Figure 2.37). Oxygen-sensitive microelectrodes (tip diameter of 5 m) to measure the distribution of PO2 in dermal papillae of the fingernail folds of healthy human subjects have been developed by Wang et al. (128).
The vasculature in the nail bed is unique in that it must supply a vascular structure between two hard surfaces, the nail plate and the bone. Studies with scanning electron microscopy revealed special vascular patterns of nail microcirculation (129). In the eponychium and perionychium, the vascular villi followed the direction of nail growth. In the face of the eponychium in contact with the nail, a wide-mesh
P.49
net of capillaries was evident. In the nail bed, the vessels were arranged in many longitudinal trabeculae parallel to the major axis of the digit. In the root of the nail, many columnar vessels characterized by multiple angiogenic buttons on their surface may be observed.
 |
Figure 2.36 Vascular system of the nail bed. This zone has a rich vascular supply, with numerous vertical arteries (branches of the arched arteries of the deep dermis) (MF stain, 47). |
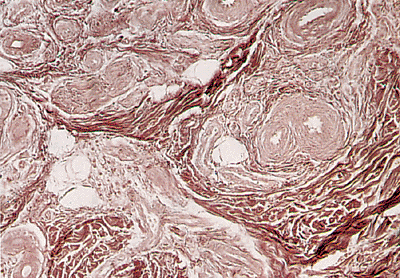 |
Figure 2.37 Detail of the rich vascular supply of the nail bed (reticulin stain, 158). |
The venous drainage is achieved by two veins, one on each side of the nail plate, in the proximal nail folds (127). The capillary network is easily seen in the proximal nail fold with a magnifying lens and is seen in more detail with an ophthalmoscopic or capillary microscope. lt is essentially the same as the network of the skin, but the capillary loops are more horizontal and visible throughout their length. Certain diseases (e.g., connective tissue disorders, macroglobulinemia, cryoglobulinemia, psoriasis, antiphospolipid syndrome) can modify its normal structure and a simple clinical examination or use of widefield nailfold microscopy can be very useful as an aid in diagnosis (130,131,132,133). A special vascular formation is present in the distal zone of the nail bed: the glomus bodies. This vascular structure has the mission of regulating the peripheral temperature by means of arteriovenous shunts (134).
Nail Growth
The rate of growth of the nail plate has been studied extensively. Normal nail growth varies between 0.1 and 1.12 mm per day, or 1.9 to 4.4 mm per month (135,136). This growth, however, is not the same in all fingers or toes. For example, fingernails grow faster than toenails. Whereas a normal fingernail grows out completely in approximately six months, a normal toenail takes 12 to 18 months to do the same (83). Nails grow faster when regenerating after avulsion (39).
Several physiological circumstances can cause variations in the nail growth (Table 2.6). Nail growth is quicker in males (39), during the day, during pregnancy (137), in persons who bite their nails (43), and in summer or warm climates (138). Conversely, nails grow more slowly in females, during the night, in toes, in winter, after age 20 (139), and during lactation (137).
Nail growth is also altered in several diseases (Table 2.6) (140,141,142,143,144,145). Nails grow quicker with psoriasis (140), pityriasis rubra pilaris (94), etretinate treatment (141), and hyperthyroidism (39); nails grow slower in cases of immobilization or paralysis (142), local ischemic conditions (135), cytostatic therapy (39,83), denutrition (143), hypothyroidism (139), and yellow nail syndrome (144). In the case of a sudden decrease in nail growth, for example, in acute infections (136), a transverse band will appear afterward, depressed in the proximal line called Beau's line. However, patients with unilateral toenail onychomycosis did not support the hypothesis that slow nail growth rate is a predisposing factor for this onychomycosis (145). The absence of K1 and K10 from healthy nail bed may also be related to the adherence of the overlying nail. In simple terms, the nail plate might be interpreted as the suprabasal layer of the nail bed, containing keratins not produced by the nail bed but affording barrier function or some other properties associated with K1, K10, and Ha1. The presence of these overlying keratins might be responsible for the lack of a granular layer and associated absence of K1 and K10. In diseases such as onychomycosis and psoriasis, where nail plate adherence is lost, a granular layer forms alongside expression of K1 and K10. (99,100,101,145)
The rate of growth of the nail plate is determined by the turnover rate of the matrix cells. Shortly after death, matrix cells do not incorporate tritiated thymidine in their nuclei; the cells appear to be incapable of DNA synthesis and cell division, and, therefore, the nail does not grow (88). Previous reports of nail growth after death are, in fact, erroneous. Apparent growth, caused by severe postmortem drying and shrinking of the soft tissues around the nail plate, is what was observed (146).
The question of where the nail plate is formed is still controversial (147). The first theories at the beginning of the century pointed toward a complete formation by the matrix (148). Years later, Lewis (8), however, concluded that the nail plate was the product of three different matrices on the basis of staining of the nail plate with a silver-protein stain and the morphology of keratinizing cells. Lewis's hypothesis was supported by differential staining of the nail plate (8), by differential interference contrast microscopy (69,70,71,72), and by ultrastructural observation of keratohyalin granules in embryonic nail (69). Lewis's hypothesis, however, has been extensively reviewed. Zaias and Alvarez (88) used radioautography to show that the nail plate was formed exclusively by the matrix in normal conditions; Samman (149) and Norton (147) confirmed it by following the incorporation of H-labeled glycine and thymidine in
P.50
human toenails; and Caputo and Dadati (46) reported that, ultrastructurally, the nail plate was a homogeneous structure with no evidence of formation from three different matrices. To add one final bit to the confusion, Samman (149) suggests that, although under normal conditions the nail plate is made exclusively by the matrix, in certain pathologic conditions the nail bed adds a ventral nail to the undersurface of the nail plate. Finally, Kato (150) published a case with an ectopic nail at the palmar tip, with a vertical growth. In this case, the proximal nail fold promotes upward growth of the nail plate in absence of a proper nail bed.
Table 2.6 Physiological and Pathological Variations that Influence Nail Growth (Sd, Syndrome) | ||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||||||||||||||||||||||||||||||||||||||
Some recent authors believe that the nail bed epithelium produces a significant 20% portion of the nail plate and thus include the nail bed in the generative portion of the nail (151). Interestingly, indirect immunofluorescence studies in the fetal nail by means of pan-anti-type I and II hair keratin antibodies revealed a positive staining in a broad band of suprabasal cells of the nail bed epithelium. However, exclusion of hair keratins hHa1, hHb5, hHb1, hHb6 and hHa4 from the adult nail bed epithelium clearly identifies the matrix as the sole origin of the nail plate (152). It has previously been speculated that a few so-called horn cells may be added by the nail bed epithelium to the underside of the nail plate. Although our study confirmed the occurrence of clearly visible K5/17-positive nail bed cells in the lower nail plate, we believe that these cells represent sectioning artefacts. Our transverse nail sections, stained for either K5/17 or hHb5, show that the boundary between the nail bed epithelium and the nail plate is extremely undulated and that the resulting narrow-spaced folds and ridges exhibit a distinctly varying height. Therefore, it is evident that already slight deviations from a vertical angle of section through the nail bed region may reveal K5-positive cells in the nail plate, which in reality stem from the tip of an adjacent epithelial fold of the bed epithelium. Collectively these data emphasize that the nail bed epithelium does not actively contribute to the formation of the nail plate. (Figure 2.38).
An important controversy is why nails grow out instead of up. Kligman (153,154) postulated that the cul-de-sac of the proximal nail groove forced the cells of the matrix to grow out. To confirm his theory he transplanted nail matrix to the forearm, producing a vertical cylinder of hard keratin that had histologic characteristies of the nail. Hashimoto et al. (69) stated later that the long axis of matrix cells in embryonic nail was directed upward and distally.
Another important question is why the nail bed accompanies the nail plate in its growth. A well-known fact is that a hemorrhage that occurs between the plate and bed will grow forward with the plate. If the plate merely moved over the bed, the blood would not move; therefore, the upper part of the bed must move out with the plate. Some authors, such as Krantz (155), Kligman (153,154), and Zaias (84), tried to study this phenomenon in an experimental way. Of all theories, the one by Zaias is most acceptable at present: he believes that the proximal nail bed moves out, either by pressure by advancing plate or because of trauma, but that the distal nail bed and hyponychium do not move. Similarly
P.51
controversial is the existence of a thin dorsal nail plate, which has been postulated on the basis of differential staining properties of the nail plate (156); a differential distribution of phospholipid, sulfhydryl, and disulphide groups (157); and ultrastructural observations (118). Moreover, the keratin pattern of the thin dorsal nail is different from that of the lower nail in that, in addition to hair keratins as a major constituent and the epithelial keratin pairs K5/K14 and K6/K16/K17 as minor components, it also contained low amounts of K1/10 (152).
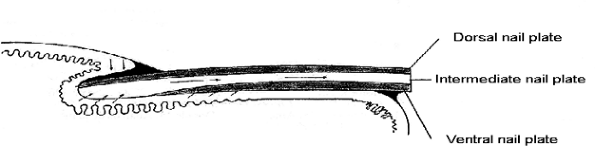 |
Figure 2.38 Schematic diagram of nail growth. |
Handling and Processing of the Nail
The major problems of processing the nail unit are the difficulty of tissue selection and the need for proper orientation of the specimen. These problems are the reason for the small number of histologic studies.
The first important point is how to take a biopsy of the nail unit (153,154,155,156,157,158,159,160,161,162,163,164,165). The best way of studying a biopsy of the nail is to ascertain that it includes the complete thickness of the nail unit (which means nail plate, bed, and subjacent dermis); these can be sectioned transversely. If a punch biopsy is taken, it should be done by boring firmly through the nail plate and into the underlying tissue to obtain a specimen with the plate attached. If one wishes to eliminate the nail plate before taking the biopsy, special care has to be taken about nail avulsion because if it has been avulsed without care, the epithelium of the bed or matrix may become separated and the undersurface may remain attached to the plate and distort the true histopathologic picture. The ideal biopsy technique for the nail is a longitudinal biopsy (159), which includes the hyponychium, the nail bed, matrix with overlying plate, and the proximal nail fold and cuticle. The specimen may be taken from the center of the nail apparatus or from the lateral edge (161,162), or it may be modified to provide elliptical excision of a tumor of the nail bed (163,164).
The second point is the orientation of the specimen for cutting. In all cases, the surgeon should alert the pathologist on the submission form as to the way the specimen was obtained, whether a particular orientation is needed, and whether a piece of the nail plate is included.
The third point is how to treat the specimen in the laboratory. If the nail plate is present in the specimen, it will be too hard for ready cutting with a microtome unless some method of softening is used. A special fixative of 5% trichloroacetic acid and 10% formalin will leave the plate softer (161). Alternatively, the specimen can be placed in distilled water for a few hours before placing in formalin (162). Another helpful technique for softening the hard keratin of the nail plate was proposed by Luna (165):
Fix specimen in 10% formalin for 24 hours.
Place specimens in the following solution until completely dekeratinized. Change solution every 2 days for best results.
- Mercuric chloride
4.0 g - Chromic acid
0.5 g - Nitric acid, concentrated
10.0 mL - Ethyl alcohol, 95%
50.0 mL - Distilled water
200.0 mL Wash in running water for three hours.
Dehydrate, clear, and impregnate with paraffin, or process as desired.
With this technique, the preservation of the cytologic characteristics of the epithelial cells of the nail matrix and nail bed is not as good as with the previous one, but it is very helpful for softening the hardest nail specimens.
References
1. Zander R. Untersuchungen uber den verhornungsprogress: 1. Die histogenese des Nagels beim menschiichen fetus. Arch Anat Entwicklungsmech 1886;273.
2. Kolliker A. Die entwicklung des menschlichen Nagels. Z Wiss Zool 1988;1:1 12.
3. Unna PG. Entwicklungsgeschichte und anatomie. In: von Ziemssen, HW. Handbuch der Speciellen Pathologie und Therapie. Vol. 14. Leipzig: 1883.
4. Boas JEV. Ein Beitrag zur Morphologie der N gel, Krallen, Hufe und Klauen der S ugetiere. Morphol Jb 1883;9:389 399.
5. Henle J. Das Wachstum des Menschlichen Nagels und des Pferdehufs. G ttingen: Dieterich; 1884.
6. Branca A. Notes sur la structure de I'ongle. Ann Dermatol Syphiligr (Paris) 1910; 1:353 371.
7. Clark WE, Buxton LH. Studies in nail growth. Br J Dermatol 1938;50:221.
8. Lewis BL. Microscopic studies of fetal and mature nail and surrounding soft tissue. AMA Arch Dermatol Syphilol 1954;70:733 747.
9. Zaias N. Embryology of the human nail. Arch Dermatol 1963;87:37 53.
10. Holbrook KA, Odland GF. The fine structure of the developing human epidermis: light, scanning, and transmission electron microscopy of the periderm. J Invest Dermatol 1975;65:16 38.
11. Conejo-Mir JS, Ambrosiani J, Dorado M. Analisis de la morfog nesis ungueal. Estudio con Microscopio Electronico de Barrido en el Embrion Humano. Barcelona: lsdin, 1985;1 8.
P.52
12. Conejo-Mir JS, Ambrosiani J, Dorado M, Camacho F, Genis BJM. Human Nail Development: A Scanning Electron Microscopy Study. Abstract book of the Meeting of the American Society of Dermatopathology. Washington, DC; 1988.
13. Suchard R. Des modifications des cellules de la matrice et du lit de l'ongle dans quelques cas pathologiques. Arch Physiol (Paris) 1882;2:445.
14. Ogura R, Knox JM, Griffin AC, Kusuhara M: The concentration of sulfhydryl and disulfide in human epidermis, hair and nail. J Invest Dermatol 1962;38:69 75.
15. Irvine AD, McLean WH. Human keratin diseases: the increasing spectrum of disease and subtlety of phenotype-genotype correlation. Br J Dermatol 1999;140,815 828.
16. De Berker D, Wojnarowska F, Sviland L, Westgate GE, Dawber RP, Leigh IM: Keratin expression in the normal nail unit: markers of regional differentiation. Br J Dermatol 2000;142:89 96.
17. Hesse M, Magin TM, Weber K. Genes for intermediate filament proteins and the draft sequence of the human genome: novel keratin genes and a surprisingly high number of pseudogenes related to keratin genes 8 and 18. J Cell Sci 2001;114,2569 2575.
18. Smith TA, Strelkov SV, Burkhard P, Aebi U, Parry DAD: Sequence comparisons of intermediate filament chains: evidence of a unique functional/structural role for coiled-coil segment 1A and linker L1. J Struct Biol 2002;137:128 145.
19. Fuchs E. Keratins and the skin. Annu Rev Cell Dev Biol 1995; 11:123 153.
20. Coulombe PA, Omary MB. Hard and soft principles defining the structure, function and regulation of keratin intermediate filaments. Curr Opin Cell Biol 2002;14:110 122.
21. Wong P, Coulombe PA. Loss of keratin 6 (K6) proteins reveals a function for intermediate filaments during wound repair. J Cell Biol 2003;163:327 337.
22. McGowan KM, Coulombe PA. Onset of keratin 17 expression coincides with the definition of major epithelial lineages during skin development. J Cell Biol 1998;143:469 486.
23. Wang Z, Wong P, Langbein L, Schweizer J, Coulombe PA: The type II epithelial keratin 6 hf (K6hf) is expressed in the companion layer, matrix and medulla of anagen-stage hair follicles. J Invest Dermatol 2003;121:1276 1283.
24. Smith FJD. Keratin 17 defect in late-onset pachyonychia. J Invest Dermatol 2004;122:268 271.
25. Ding YG, Zhu TS, Jiang W, et al. Identification of a novel mutation and a de novo mutation in DKC1 in two Chinese pedigrees with dyskeratosis congenita. J Invest Dermatol 2004;123:470 473.
26. Bowden PE, Haley JL, Kansky A, Rothnagel JA, Jones DO, Turner RJ: Mutation of a type II keratin gene (K6a) in pachyonychia congenita. Nature Genet 1995;10:363 365.
27. Smith FJF, Jonkman MF, van Goor H, et al. A mutation in human keratin K6b produces a phenocopy of the K17 disorder pachyonychia congenita type 2. Human Mol Genet 1998;7:1143 1148.
28. Available at: http://www.interfil.org. Accessed January 25, 2006.
29. Available at: http://www.genome.cse.ucsc.edu/. Accessed January 25, 2006.
30. Available at: http://www.marshmed.org/genetics/. Accessed January 25, 2006.
31. Baran R, Dawber RP, Haneke E. Hair and nail relationship. Skinmed 2005;4:18 23.
32. De Berker DA, Perrin C, Baran R. Localized longitudinal erythronychia: diagnostic significance and physical explanation. Arch Dermatol 2004;140:1253 1257.
33. Norgett EE, Wolf F, Balme B, et al. Hereditary white nails : a genetic and structural study. Br J Dermatol 2004;151:65 72.
34. Lewin K. The normal fingernail. Br J Dermatol 1965;77:421 430.
35. Pinkus F. The development of the integument. In: Keibel F, Mall F, eds. Manual of Human Embryology. Philadelphia: Lippincott; 1910:243 291.
36. Schweitzer TP, Rayan GM. The terminal tendon of the digital extensor mechanism: Part I, anatomic study. J Hand Surg Am 2004;29:898 902.
37. Le Gros Clark WB. The problems of the claw in primates. Proc Zool Soc Lond 1936;I:1 24.
38. Pinkus F. Der Nagel. In: Jadassohns Handbuch, der Haut- und Geschlechtskrankheiten. Berlin:Springer-Verlag; 1927.
39. Baran R, Dawber RPR, eds. Diseases of the Nail and Their Management. Oxford: Blackwell Scientific; 1984:1 21.
40. Caputo R, Cappio F, Rigoni C, et al. Pterygium inversum unguis. Report of 19 cases and review of the literature. Arch Dermatol 1993;129:1307 1309.
41. Terry RB. The onychodermal hand in health and disease. Lancet 1955;1:179 18l.
42. Martin BF, Platts MM. A histological study of the nail region in normal human subjects and in those showing splinter hemorrhages of the nail. J Anat 1959;93:323 330.
43. Raffle EJ. Terry's nails. Lancet 1984;1:1131.
44. Finlay AY, Moseley H, Duggan TC. Ultrasound transmission time: an in vivo guide to nail thickness. Br J Dermatol 1987;117:765 770.
45. Goettmann S, Drape JL, Idy-Peretti I, et al. Magnetic resonance imaging: a new tool in the diagnosis of tumors of the nail apparatus. Br J Dermatol 1994;130:701 710.
46. Caputo R, Dadati E. Preliminary observations about the ultrastructure of the human nail plate treated with thioglycolic acid. Arch Klin Exp Dermatol 1968; 231:344 354.
47. Forslind B, Thyresson N. On the structure of the normal nail. A scanning electron microscope study. Arch Dermatol Forsch 1975;251:199 204.
48. Garson JC, Baltenneck F, Leroy F, et al. Histological structure of human nail as studied by synchrotron X-ray microdiffraction. Cell Mol Biol 2000;46:1025 1034.
49. Hamilton JB, Terada H, Mestler GE. Studies of growth throughout the lifespan in Japanese. Growth and size of nails and their relationship to age, sex, hereditary and other factors. J Gerontol 1955;10:401 415.
50. Johnson M, Shuster S. Determinants of nail thickness and length. Br J Dermatol 1994;130:195 198.
51. Farren L, Shayler S, Ennos AR. The fracture properties and mechanical design of human fingernails. J Exp Biol 2004;207 (pt 5):735 741.
52. Germann H, Barran W, Plewig G. Morphology of corneocytes from human nail plates. J Invest Dermatol 1980;74:115 118.
53. Walters KA, Flynn GL, Marvel JR. Physicochemical characterization of the human nail: I. Pressure sealed apparatus for measuring nail plate permeabilities. J Invest Dermatol 1981;76:76 79.
54. Egawa M, Fukuhara T, Takahashi M, Ozaki Y. Determining water content in human nails with a portable near-infrared spectrometer. Appl Spectrosc 2003;57:473 478.
55. Nuutinen J, Harvima I, Lahtinen MR, Lahtinen RT. Water loss through the lip, nail, eyelid skin, scalp skin and axillary skin measured with a closed-chamber evaporation principle. Br J Dermatol 2003;148:839 842.
56. Jarrett A, Spearman RI. The histochemistry of the human nail. Arch Dermatol 1966;94:652 657.
57. Pautard FGE. Mineralization of keratin and its comparison with the enamel matrix. Nature 1963;199:531 540.
58. Forslind B, Wroblewski R, Afzelius BA. Calcium and sulphur location in human nail. J Invest Dermatol 1976;67:273 290.
59. Helmdach M, Thielitz A, Ropke EM, Gollnick H. Age and sex variation in lipid composition of human fingernail plates. Skin Pharmacol Appl Skin Physiol 2000;13:111 119.
60. Sass JO, Skladal D, Zelger B, Romani N, Utermann B. Trichothiodystrophy: quantification of cysteine in human hair and nails by application of sodium azide-dependent oxidation to cysteic acid. Arch Dermatol Res 2004;296:188 191.
61. Bolliger A, Gross R. Non-keratin of human toenails. Aust J Exp Biol Med Sci 1953;127 130.
62. Levitt JI. Creatinine concentration of human fingernail and toenail clippings. Application in determining the duration of renal failure. Ann Intern Med 1966;64:312 327.
63. Goldblum RW, Derby S, Lerner AB. The metal content of skin, nails and hair. J Invest Dermatol 1953;20:13 18.
64. Kopito L, Mahmoodian A, Townley RRW, Khaw KT, Shwachman H. Studies in cystic fibrosis: analysis of nail clippings for sodium and potassium. N Engl J Med 1965;272:504 509.
65. Martin GM. Copper content of hair and nails of normal individuals and of patients with hepatolenticular degeneration. Nature 1964;202:903 904.
66. Mandal BK, Ogra Y, Anzai K, Suzuki KT. Speciation of arsenic in biological samples. Toxicol Appl Pharmacol 2004;198:307 318.
P.53
67. Cingolani M, Scavella S, Mencarelli R, Mirtella D, Froldi R, Rodriguez D. Simultaneous detection and quantitation of morphine, 6-acetylmorphine, and cocaine in toenails: comparison with hair analysis. J Anal Toxicol 2004;28:128 131.
68. Gillespie JM, Frenkel MJ. The diversity of keratins. Comp Biochem Physiol 1974;47B:339 346.
69. Hashimoto K, Gross BG, Nelson R, Lever WF. The ultrastructure of the skin of human embryos. III. The formation of the nail in 16 18-week-old embryos. J Invest Dermatol 1966;47:205 217.
70. Hashimoto K. Ultrastructure of the human toenail. l. Proximal nail matrix. J Invest Dermatol 1971;56:235 246.
71. Hashimoto K. Ultrastructure of the human toenail. II. Keratinization and formation of the marginal band. J Ultrastruct Res 1971;36:391 410.
72. Hashimoto K. Ultrastructure of the human toenail. III. Cell migration, keratinization and formation of the intercellular cement. Arch Dermatol Forsch 1971;240:1 22.
73. Higashi N. Melanocytes of nail matrix and nail pigmentation. Arch Dermatol 1968;97:570 574.
74. Scott GA, Haake AR. Keratinocytes regulate melanocyte number in human fetal and neonatal skin equivalents. J Invest Dermatol 1991;97:776 781.
75. Higashi N, Saito T. Horizontal distribution of dopa-positive melanocytes in the nail matrix. J Invest Dermatol 1969;53:163 165.
76. Jimbow K, Takahashi M, Sato S, Kukita A. Ultrastructural and cytochemical studies on melanogenesis in melanocytes of normal human hair matrix. J Electron Microsc (Tokyo) 1971;20:87 92.
77. Feibleman CE, Stoll H, Maize JC. Melanomas of the palm, sole and nail bed: a clinicopathologic study. Cancer 1980;46:2492 2504.
78. Baran R, Juhlin L. Photoonycholysis. Photodermatol Photoinmunol Photomed 2002;18:202 208.
79. Gallais V, Lacour JP, Perrin C, Ghanem G, Bodokh I, Ortonne JP. Acral hyperpigmented macules and longitudinal melanonychia in AIDS patients. Br J Dermatol 1992;126:387 391.
80. Uyttendaele H, Geyer A, Scher RK. Drugs and nails. J Eur Acad Dermatol Venereol 2004;18:124 125.
81. Moll I, Moll R. Merkel cells in ontogenesis of human nails. Arch Dermatol Res 1993;285:366 371.
82. Samman PD. The ventral nail. Arch Dermatol 1961;84:1030 1033.
83. Samman PD. The Nails in Disease. 3rd ed. London: Heinemann Medical Books; 1978.
84. Zaias N. The movement of the nail bed. J Invest Dermatol 1967; 48:402 403.
85. Drape JL, Wolfram-Gabel W, Idy-Peretti I, et al. The lunula: a magnetic resonance imaging approach to the subnail matrix area. J Invest Dermatol 1996;106:1081 1085.
86. Dawber RPR, Baran R. The nails. In: Rook A, Wilkinson DS, Ebling FJG, Champion RH, Burton JL, eds. Textbook of Dermatology. Oxford: Blackwell Scientific; 1986:2039 2044.
87. Burrows MT. The significance of the lunula of the nail. Johns Hopkins Hosp Res 1919;18:357 361.
88. Zaias N, Alvarez J. The formation of the primate nail plate. An autoradiographic study in squirrel monkeys. J Invest Dermatol 1968;51:120 136.
89. Meyer JC, Grundmann HP. Scanning electron microscopic investigation of the healthy nail and its surrounding tissue. J Cutan Pathol 1984;11:74 79.
90. Omura EF. Histopathology of the nail. Dermatol Clin 1985; 3:531 541.
91. Sinclair RD, Wojnarowska F, Leigh IM, Dawber RP. The basement membrane zone of the nail. Br J Dermatol 1994;131:499 505.
92. Dorschner RA, Lopez-Garcia B, Massie J, Kim C, Gallo RL. Innate immune defense of the nail unit by antimicrobial peptides. J Am Acad Dermatol 2004;50:343 348.
93. Holbrook KA. Human epidermal embryogenesis. Int J Dermatol 1979;18:329 356.
94. Runne U, Orfanos CE. The human nail: structure, growth and pathological changes. Curr Probl Dermatol 1981;9:102 149.
95. Zaias N. Onychomycosis. Arch Dermatol 1972;105:263 274.
96. Baden HP, Kvedar JC. Epithelial cornified envelope precursors are in the hair follicle and nail. J Invest Dermatol 1993; 101(suppl1):72S 74S.
97. O'Keefe EJ, Hamilton EH, Lee SC, Steinert P. Trichohyalin: a structural protein of hair, tongue, nail and epidermis. J Invest Dermatol 1993;101(suppl 1):65S 71S.
98. Heid HW, Moll I, Franke WW. Pattern of expression of trichocytic and epithelial cytokeratins in mammalian tissues. II. Concomitant and mutually exclusive synthesis of trichocytic and epithelial cytokeratins in diverse human and bovine tissues (hair follicle, nail bed and matrix, lingual papilla, thymic reticulum). Differentiation 1988;37:215 230.
99. Kitahara T, Ogawa H. The extraction and characterization of human nail keratin. J Dermatol Sci 1991;2:402 406.
100. Kitahara T, Ogawa H. Cultured nail keratinocytes express hard keratins characteristic of nail and hair in vivo. Arch Dermatol Res 1992;284:253 256.
101. Kitahara T, Ogawa H. Coexpression of keratins characteristic of skin and hair differentiation in nail cells. J Invest Dermatol 1993;100:171 175.
102. De Berker D, Wojnarowska F, Sviland L, Westgate GE, Dawber RPR, Leigh IM. Keratin expression in the normal nail unit: markers of regional differentiation. Br J Dermatol 2000;142:89 96.
103. Heid HW, Moll I, Franke WW. Patterns of expression of trichocytic and epithelial cytokeratins in mammalian tissues. II. Concomitant and mutually exclusive synthesis of trichocytic and epithelial cytokeratins in diverse human and bovine tissues (hair follicle, nail bed and matrix, lingual papilla, thymic reticulum). Differentiation 1988;37:215 230.
104. Perrin C, Langbein L, Schweizer J. Expression of hair keratins in the adult nail unit: an immunohistochemical analysis of the onychogenesis in the proximal nail fold, matrix and nail bed. Br J Dermatol 2004;151:362 371.
105. Kitahara T, Ogawa H. Cellular features of differentiation in the nail. Microsc Res Tech 1997;38:436 442.
106. Lane EB, Wilson CA, Hughes BR, Leigh IM. Stem cells in hair follicles. Cytoskeletal studies. Ann NY Acad Sci 1991;642:197 213.
107. Stark HJ, Breitkreutz D, Limat A, et al. Keratins of the human hair follicle: hyperproliferative keratins consistently expressed in outer root sheath cells in vivo and in vitro. Differentiation 1987;35:236 248.
108. McLean WH Irwin, Epithelial Genetics Group. Genetic disorders of palm skin and nail. J Anat 2003;202:133 141.
109. Inoue S, Nambu T, Shimomura T. The RAIG family member, GPRC5D, is associated with hard-keratinized structures. J Invest Dermatol 2004;122:565 573.
110. Okazaki M, Yoshimura K, Fujiwara H, Suzuki Y, Harii K. Induction of hard keratin expression in non-nail-matrical keratinocytes by nail-matrical fibroblasts through epithelial-mesenchymal interactions. Plast Reconstr Surg 2003;111:286 290.
111. Egawa K, Kuroki M, Inoue Y, Ono T. Nail bed keratinocytes express an antigen of the carcinoembryonic antigen family. Br J Dermatol 2000;143:79 83.
112. Lavker RM, Risse B, Brown H, et al. Localization of plasminogen activator inhibitor type 2 (PAI-2) in hair and nail: implications for terminal differentiation. J Invest Dermatol 1998;110:917 922.
113. Lacour JP, Dubois D, Pisani A, Ortonne JP. Anatomical mapping of Merkel cells in normal human adult epidermis. Br J Dermatol 1991;125:535 542.
114. Tosti A, Cameli N, Piraccini BM, Fanti PA, Ortonne JP. Characterization of nail matrix melanocytes with anti-PEP1, anti-PEP8, TMH-1 and HMB-45 antibodies. J Am Acad Dermatol 1994;31(pt 1):193 196.
115. Guerrero-Fernandez J, Garcia-Ascaso MT, Guerrero Vazquez J. Pigmentation band on toenail [in Spanish]. An Pediatr (Barc). 2004;61:455 456.
116. Cameli N, Picardo M, Tosti A, Perrin C, Pisani A, Ortonne JP. Expression of integrins in human nail matrix. Br J Dermatol 1994;130:583 588.
117. Picardo M, Tosti A, Marchese C, et al. Characterization of cultured nail matrix cells. J Am Acad Dermatol 1994;30:434 440.
118. Doigel AS. Die nerbenendigungen im nagelbett des Menschen. Arch Mikros Anat 1904;64:173 188.
P.54
119. Martino L. Sulia innervazione dell'apparato ungueale. Boll Soc Ital Biol Sper 1942;1;7:488 489.
120. Winkelmann RK. Nerve Endings in Normal and Pathologic Skin. Springfield, IL: Charles C Thomas; 1960:100.
121. Reginelli AD, Wang YQ, Sassoon D, Muneoka K. Digit tip regeneration correlates with regions of Msx1 (Hox7) expression in fetal and newborn mice. Development 1995;121:1065 1076.
122. Mohammad KS, Neufeld DA. Denervation retards but does not prevent toetip regeneration. Wound Repair Regen 2000;8:277 281.
123. Zhao W, Neufeld DA. Bone regrowth in young mice stimulated by nail organ. J Exp Zool 1995;271:155 159.
124. Flint MH. Some observations on the vascular supply of the nail bed and terminal segments of the finger. Br J Plastic Surg 1955;8:186 195.
125. Ryan TJ. The blood vessels of the skin. In: Jarret A, ed. The Physiology and Pathophysiology of the Skin. Vol 2. London: Academic Press; 1973:612:658 659.
126. Smith DO, Oura C, Kimura C, Toshimori K. Artery anatomy and tortuosity in the distal finger. J Hand Surg Am 1991;16:297 302.
127. Hale AR, Burch GE. The arteriovenous anastomoses and blood vessels of the human finger. Medicine (Baltimore) 1960;39:191 240.
128. Wang W, Winlove CP, Michel CC. Oxygen partial pressure in outer layers of skin of human finger nail folds. J Physiol 2003;549:855 863.
129. Sangiorgi S, Manelli A, Congiu T, et al. Microvascularization of the human digit as studied by corrosion casting. J Anat 2004; 204:123 131.
130. Ross JB. Nail fold capillaroscopy: a useful aid in the diagnosis of collagen vascular diseases. J Invest Dermatol 1966;47:282 285.
131. Gilje O, Kierland R, Baldes EJ. Capillary microscopy in the diagnosis of dermatologic diseases. J Invest Dermatol 1974;22:199 206.
132. Ohtsuka T, Yamakage A, Miyachi Y. Statistical definition of nailfold capillary pattern in patients with psoriasis. Int J Dermatol 1994;33:779 782.
133. Vaz JL, Dancour MA, Bottino DA, Bouskela E. Nailfold videocapillaroscopy in primary antiphospholipid syndrome (PAPS). Rheumatology 2004;43:1025 1027.
134. Mehregan AH. Pinkus' Guide to Dermatohistopathology. 4th ed. Norwalk, CT: Appleton-Century-Crofts; 1986:563 564.
135. Bean WB. Nail growth: 30 years of observation. Arch Intern Med 1974;134:497 502.
136. Sibinga MS. Observations on growth of fingernails in health and disease. Pediatrics 1959;24:225 233.
137. Halban J, Spitzer MZ. On the increased growth of nails in pregnancy. Monatsschr Gerburtshilfe Gynaekol 1929;82:25.
138. Geoghegan B, Roberts DF, Sampford MR. A possible climatic effect on nail growth. J Appl Physiol 1958;13:135 138.
139. Orentreich N, Markofsky J, Vogelman JH. The effect of aging on the rate of linear nail growth. J Invest Dermatol 1979;73:126 130.
140. Landherr G, Braun-Falco O, Hofmann C, Plewig G, Galosl A. Fingernagelwachstum bei Psoriatikem unter puvatherapie. Hautarzt 1982;33:210 213.
141. Baran R. Action th rapeutique et complications due r tino de sur l'appareil ungu al. Ann Dermatol V n r ol 1982;109:367 371.
142. Fleckman P. Anatomy and physiology of the nail. Dermatol Clin 1985;3:373 381.
143. Geyer AS, Onumah N, Uyttendaele H, Scher RK. Modulation of linear nail growth to treat disease of the nail. J Am Acad Dermatol 2004;50:229 234.
144. Pavlidakey GP, Hashimoto K, Blum D. Yellow nail syndrome. J Am Acad Dermatol 1984;11:509 512.
145. Yu HJ, Kwon HM, Oh DH, Kim JS. Is slow nail growth a risk factor for onychomycosis? Clin Exp Dermatol 2004;29:415 418.
146. Zaias N. Nails. components, growth and composition of the nail. In: Demis DJ, Dobson RL, McGuire J, eds. Clinical Dermatology. Vol. 1. New York: Harper & Row; 1980:1 6.
147. Norton LA. Incorporation of thymidine-methyl-H3 and glycine-2-H3 in the nail matrix and bed of humans. J Invest Dermatol 1971;56:61 68.
148. Unna PG. Entwicklungsgeschichte und anatomie. In: von Ziemssen, HW Handbuch der Speciellen Pathologie und Therapie. Vol 14. Leipzig: 1883.
149. Samman PD. The human toe nail. Its genesis and blood supply. Br J Dermatol 1959;71:296 302.
150. Kato N. Vertically growing ectopic nail. J Cutan Pathol 1992; 19:445 447.
151. Johnson M, Comaish JS, Shuster S. Nail is produced by the normal nail bed: a controversy resolved. Br J Dermatol 1991;125:27 29.
152. De Berker D, Mawhinney B, Sviland L. Quantification of regional matrix nail production. Br J Dermatol 1996;134:1083 1086.
153. Kligman AM. Nails. In: Pillsbury DM, Shelley WB, Kligman AM, eds. Dermatology. Philadelphia: WB Saunders; 1956:80 86.
154. Kligman AM. Why do nails grow out instead of up? Arch Dermatol 1961;84:313 315.
155. Krantz W. Beitrag zur anatomie des nagels. Dermatol Z 1937; 239 242.
156. Achten G, Andre J, Laporte M. Nails in light and electron microscopy. Semin Dermatol 1991;10:54 64.
157. Lynch MH, O'Guin WM, Hardy C, Mak L, Sun TT. Acidic and basic hair/nail ( hard ) keratins: their colocalization in upper cortical and cuticle cells of the human hair follicle and their relationship to soft keratins. J Cell Biol 1986;103(pt 2): 2593 2606.
158. Parent D, Achten G, Stoofs-Vamhoof F. Ultrastructure of the normal human nail. Am J Dermatopathol 1985;7:529 535.
159. Baran R, Sayag J. Nail biopsy: why, when, where, how? J Dermatol Surg 1976;2:322 324.
160. Bennet RG. Technique of biopsy of nails. J Dermatol Surg 1976;2:325 326.
161. Stone OJ, Barr RJ, Herten RJ. Biopsy of the nail area. Cutis 1978; 21:257 260.
162. Scher RK. Biopsy of the matrix of a nail. J Dermatol Surg Oncol 1980;6:19 21.
163. Scher RK. Longitudinal resection of nails for purposes of biopsy and treatment. J Dermatol Surg Oncol 1980;6:805 807.
164. Rich P. Nail biopsy: indications and methods. J Dermatol Surg Oncol 1992;18:673 682.
165. Luna LG. Preparation of tissues. In: Luna LG. Manual of Histologic Staining Methods of the Armed Forces Institute of Pathology. 3rd ed. New York: McGraw-Hill Book Company; 1968:1 12.