18 - Lungs
Editors: Mills, Stacey E.
Title: Histology for Pathologists, 3rd Edition
Copyright 2007 Lippincott Williams & Wilkins
> Table of Contents > VII - Alimentary Tract > 24 - Small Intestine
function show_scrollbar() {}
24
Small Intestine
Terry L. Gramlich
Robert E. Petras
Gross Anatomy and Surgical Perspective
The small intestine, located within the abdominal cavity, is a multiply coiled tubular organ that extends from the gastric pylorus to the junction of the cecum and ascending colon. Its average length in the human adult is 6 to 7 m (1). Three subdivisions the duodenum, jejunum, and ileum are defined and characterized by various anatomic relationships. The duodenum is the most proximal portion of the small intestine; it measures about 12 inches (20 25 cm) in length and extends from the pylorus to the duodenojejunal flexure. The duodenum, excluding the most proximal several centimeters, is a fixed, retroperitoneal structure that forms a C- or U-shape around the head of the pancreas (2). Four subdivisions of the duodenum have been described: (a) the first portion, also known as the duodenal cap or bulb, is the most proximal and superior segment; (b) the descending or second portion, into which the common bile duct and major and minor pancreatic ducts empty into their respective papillae; (c) the horizontal or third portion; and (d) the ascending or fourth portion, which veers forward at the level of the second lumbar vertebra, just left of midline, to become continuous with the remainder of the small bowel (1,2).
The origin of the jejunum is marked by a strip of fibromuscular tissue, the so-called ligament of Treitz, which anchors the terminal duodenum and duodenojejunal flexure to the posterior abdominal wall (3). Distal to the ligament of Treitz, the remainder of the small bowel is arbitrarily subdivided into the jejunum (the proximal two-fifths) and the ileum (the distal three-fifths, terminating at the ileocecal junction within the right iliac fossa) (1).
Although a discrete point demarcating jejunum from ileum does not exist, several relatively distinctive features become gradually more apparent from proximal to distal; these features help surgeons isolate specific segments of the small bowel. For example, the proximal jejunum has a thicker wall
P.604
and is about twice the diameter of distal ileum. In addition, jejunal segments have more prominent permanent circular folds (plicae circulares) that can be palpated externally at surgery (1,4). The quantity of mesenteric adipose tissue is greater in the ileum, thus imparting a dense opaque appearance that contrasts with the less fatty, translucent mesentery of the jejunum (4). Finally, most of the jejunum lies within the upper abdominal cavity, whereas most of the ileum lies within the lower abdominal cavity and pelvis.
The arterial vascular supply of the small bowel originates from two major aortic axes: the celiac and superior mesenteric trunks (5). The duodenum is supplied by branches and interanastomosing arcades of both trunks, and its blood supply is intimately associated with that of the pancreatic head (1). The jejunum and ileum receive their blood from more distal branches of the superior mesenteric artery (5). The lymphatic and venous drainage systems follow the arterial supply and flow into regional lymphatics and lymph nodes or the portal venous system, respectively.
Sympathetic neural input to the small bowel is carried by the celiac and superior mesenteric plexuses, whereas the parasympathetic supply is derived from distal branches of the vagus nerve; these both closely follow the arterial paths into the bowel wall.
Physiology
The small intestine has several functional roles, the most important of which is the processing and absorption of ingested nutrients. Pancreatic enzymes act on the larger ingested carbohydrates and proteins to produce more appropriately sized molecules for further digestion. The brush border created by the numerous apical microvilli on absorptive epithelial cells offers an array of peptidases and carbohydrases, which act as key enzymes in additional nutrient breakdown and processing (6). The resulting smaller molecules are subsequently absorbed across the epithelial layer, and most pass into the portal venous system for eventual systemic distribution.
Fat digestion differs somewhat in that pancreatic lipase and bile act intraluminally to create free fatty acids and monoglycerides that migrate via direct diffusion through the lipid-soluble plasma membrane of enterocytes without surface processing or active transport (7). Most undergo intracytoplasmic resynthesis with directed packaging into chylomicrons and are released into regional lymphatic vessels. Water and electrolytes, vitamins, minerals, and various drugs also are absorbed at points along the mucosa of the small bowel. Therefore, the structural integrity of this viscus is critical to the maintenance of nutritional status, as well as in appropriate drug handling.
The small intestine also functions to propel and segmentally mix both newly accepted gastric contents and the residual material left after initial digestive efforts. Although a number of factors influence gut motility, the most basic contractile activity is initiated at the level of the individual smooth muscle cells within the wall (8). Important functional differences exist, based on whether an individual is feeding or fasting. With feeding, a distended bowel segment initiates peristalsis, a forward propulsive motion that is mediated through the enteric nervous system; the intrinsic neurons of the myenteric plexus of Auerbach are most important in this regard (8,9). In contrast, during fasting or between meals, a slow yet continually recurring set of contractions attempts to clear the enteric lumen of any residual debris. The hormone motilin is believed to be important in the generation of these migratory motor complexes (9). Other endocrine influences, as well as the autonomic and central nervous systems, play a modulatory role in these intrinsic activities.
Advances in the burgeoning field of endocrine gut physiology and immunohistology have disclosed a variety of hormones within individual cells lining the small intestinal mucosa (10). Although the precise physiologic role of most of these cells and their secretory products remains to be determined, some are thought to exert a modulatory effect on gut motility or to influence the function of nearby epithelial cells (9,11).
The gut in general and the small bowel in particular have a crucial function in mucosal immunity. The mucosa/gut-associated lymphoid tissues, which are discussed in detail later in this chapter, are important in the local defense against mucosally encountered microorganisms and generate the initial immunologic responses to these various agents (12). Additionally, these tissues are the breeding ground for various reactive and neoplastic pathologic conditions.
Histology
Although regional histologic differences exist within the small intestine, the general microscopic structure is similar throughout its length. The wall of the small bowel can be divided into four basic layers: mucosa, submucosa, muscularis externa or propria, and serosa.
Mucosa
Mucosal Architecture and Design
Because the principal function of the small intestine is absorption of ingested nutrients, the mucosa, which is the layer in contact with luminal contents, is specifically designed for this purpose. Several architectural adaptations augment the otherwise limited surface area of the small intestine (13). One of these, the grossly evident permanent circular folds (plicae circulares), courses perpendicular to the longitudinal axis of the bowel (13,14) (Figure 24.1). These mucosa-covered folds contain submucosal cores and
P.605
traverse nearly the entire circumference of the bowel lumen before overlapping with adjacent permanent folds. In addition to enhancing surface area, they act as partial barriers that attenuate the forward flow of intraluminal contents, thus increasing the time of contact with absorptive surfaces.
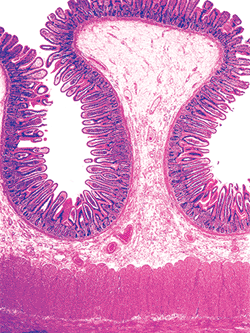 |
Figure 24.1 A single plica, or permanent circular fold, with its submucosal core and mucosal surface. The absorptive surface area is further augmented by intraluminal mucosal projections (villi). |
The mucosa is composed of an epithelial component, a lamina propria, and a muscularis mucosae. The surface epithelium and lamina propria form intraluminal projections called villi. These microscopic fingerlike and leaflike projections cover the entire luminal surface of the small bowel and are the most important morphologic modification responsible for enhancing surface area (13,15,16) (Figures 24.2,24.3). Each villous surface is covered by a single layer of epithelium consisting of various cell types. Beneath this epithelial layer lies a core of lamina propria that contains a centrally located, blind-ended lymphatic channel (lacteal), an arteriovenous capillary network, and an abundant migratory cell population (17,18,19) (Figure 24.4).
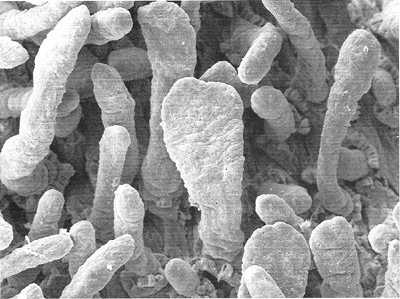 |
Figure 24.2 Scanning electron micrograph of small intestinal mucosa discloses the fingerlike and leaflike appearance of villi. Fingerlike villi predominate in the more distal segments of small bowel (jejunum and ileum), whereas leaflike villi are more common in the duodenum. Mixed populations, as in this micrograph, are considered normal. |
In the intervening regions and beneath the villi lie the crypts of Lieberk hn. These tubular intestinal glands open between the villi and extend down to the muscularis mucosae (Figure 24.3). The crypts are depressions of the surface epithelium, whereas the villi are extensions above it. However, these mucosal compartments are contiguous in that the lamina propria forming the villous cores also surrounds the crypts. The ratio of villous length to crypt length in normal small bowel varies from about 3:1 to 5:1 (13) (Figure 24.3). The crypts and surrounding lamina propria lie upon the muscularis mucosae, a thin fibromuscular layer that separates the mucosa from the underlying submucosa.
Mucosal Components and Their Composition
Epithelium
The mucosal epithelium is divided into the villous and crypt compartments. Although similar in appearance, the cell types differ somewhat, and their basic functions are distinct. Common to both, however, is a basic polarity of cellular organization, with nuclei aligned side by side, typically in a basal location within each cell.
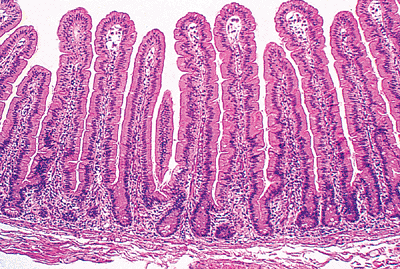 |
Figure 24.3 Normal jejunal villi. These villi are long and slender mucosal projections with a core of lamina propria covered by a luminal epithelial layer. A single row of intestinal glands (crypts) is found at the base of the mucosa. These crypts lie between adjacent villi and are surrounded by the same lamina propria that forms the villous cores. |
P.606
Villous Epithelium.
The absorptive cell is the major villous epithelial cell type encountered. It is tall, columnar, with a basally situated round-to-oval nucleus and an eosinophilic cytoplasm (Figure 24.5). The apical surface contains a brush border that appears densely eosinophilic, stains positive with periodic acid-Schiff (PAS), and is composed of microvilli and the glycocalyx, or fuzzy coat (Figures 24.5,24.6). Microvilli, which are best seen on ultrastructural examination, are evenly spaced surface projections that also augment the mucosal surface area of the small intestine (20) (Figure 24.7). Multiple filamentous structures emanating from and contiguous with their surface comprise the glycocalyx (17). The microvillous membrane glycocalyx complex houses important enzymes peptidases and disaccharidases that function in terminal digestive processes. This layer probably also acts as a barrier to microorganisms and other foreign matter (21). Components forming the glycocalyx are continually
P.607
synthesized within the absorptive cell and transported to the surface to replace the preexisting coat in a dynamic fashion (17,21,22). Although its functional capacities are uncertain, absence of the glycocalyx of the small bowel mucosa was the sole detectable histologic abnormality found in some children with allergic enteropathy (i.e., cow's milk allergy) (21).
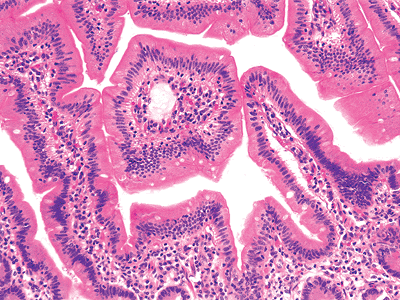 |
Figure 24.4 The duodenal villous surface is covered by a single layer of tall columnar epithelial cells. The underlying lamina propria core contains lymphoid and plasma cells and a connective tissue framework, including a lymphatic vessel (lacteal) and a subepithelial capillary network. |
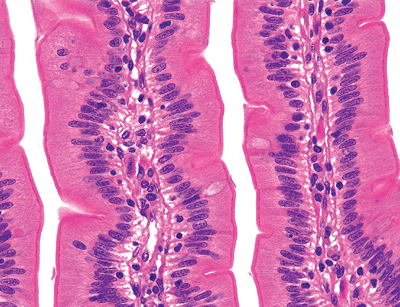 |
Figure 24.5 High-magnification view of a jejunal villi disclosing the general features of villous morphology. Both columnar absorptive cells and goblet cells (with apical clear vacuole) cover the villous surfaces; each cell type has a basally situated oval-to-round nucleus. Microvilli (brush border) are seen extending from the columnar absorptive cell surface. Note the intraepithelial lymphocytes scattered among and between the epithelial cells. |
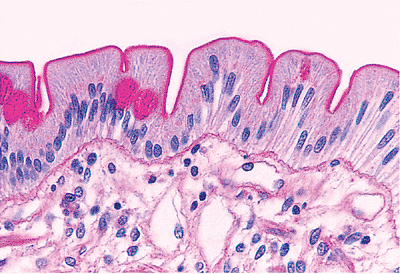 |
Figure 24.6 Periodic acid-Schiff (PAS) stain highlights the microvillous membrane glycocalyx complex along the apical surface of the absorptive cells. The thin subepithelial basement membrane that separates the lamina propria from the epithelial compartment also stains with PAS but to a lesser degree. The neutral subgroup of mucins contained within the goblet cells are PAS-positive as well. |
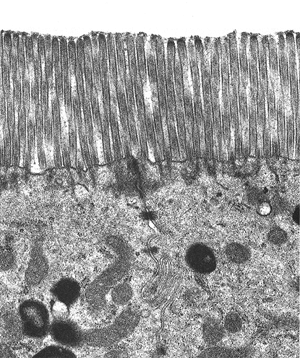 |
Figure 24.7 Transmission electron micrograph of microvilli emanating from the absorptive columnar cell surfaces. The glycocalyx component is the filamentous layer overlying the microvilli, but most of this has been artifactually removed during processing. |
Interspersed among the absorptive cells are goblet cells, which have a characteristic apical mucin droplet and an attenuated, basally situated, bland nucleus (Figure 24.5). They contain both neutral and acid mucin and function as secretory cells, sustaining a moist viscid environment within the lumen (20). In a combined Alcian blue/PAS stain, the droplets appear blue-purple (Figure 24.8). The acid mucins of the small intestine are primarily sialomucins, in contrast to the colonic goblet cell, which contains predominantly acid sulfomucins (23,24). The number of goblet cells increases with distal progression along the small bowel (20).
Scattered endocrine cells are present within the villous epithelium, but they are more abundant within the crypts. Intraepithelial lymphocytes are scattered among and lie between individual epithelial cells, usually just above the basement membrane; normally there is about one lymphocyte for every five epithelial cells (25,26) (Figure 24.5). Intraepithelial lymphocytes mark as T cells (Figure 24.9), and most are of the T-suppressor/cytotoxic (CD8-positive) variety (27,28,29). An increase in the number of intraepithelial lymphocytes is characteristic of several disorders, including gluten-sensitive enteropathy (celiac sprue), tropical sprue, and giardiasis (30,31).
Crypt Epithelium.
The crypt epithelium primarily functions in epithelial cell renewal (20); and, as a consequence of this regenerative function, mitoses are seen frequently within the crypts (normal range: 1 12 mitoses/crypt) (32). Goblet cells and columnar cells, some of which are undifferentiated or stem cells, are conspicuous. The four major epithelial cell types of the mucosa (absorptive, goblet, endocrine, and Paneth's cells) arise from this stem cell (33). Differentiation and maturation occur in about five to six days as the cells migrate from the crypt depths to the villous tips, where they are subsequently sloughed into the lumen (20,33); however, the Paneth's cell remains within the crypt base (34). It is thought that apoptosis probably functions as a regulator of cell migration toward the villous surface; however, it is not certain whether this form of programmed cell death is responsible for epithelial cell sloughing from the villous tip (35). Morphologically, cells undergoing apoptosis show nuclear condensation and cell shrinkage with subsequent detachment from adjacent epithelium. In addition,
P.608
the cell disintegrates into so-called apoptotic bodies and is phagocytosed by local histiocytes (35).
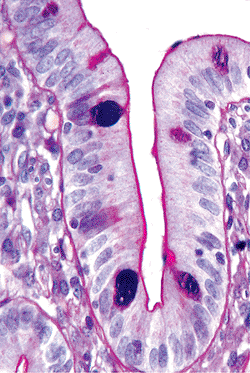 |
Figure 24.8 Alcian blue/PAS combination stain showing characteristic blue-purple apical mucin droplet of intestinal goblet cells. The heterogeneous composition (neutral and acid mucins) allows both stains to be incorporated into the droplet, imparting this distinctive color. |
Endocrine cells are relatively abundant in the crypts, occurring as single cells or in discontinuous groupings along the intestinal tract (10). They are of two morphologic types. The open type have a pyramidal shape that tapers toward the glandular lumen with which they communicate, whereas closed cells are spindle-shaped and have no luminal connection (10). The former are the most frequent type found in the small bowel. Some endocrine cells disclose eosinophilic basal (infranuclear) granules on hematoxylin and eosin (H&E) staining so that they are easily identified on routine preparations (Figure 24.10); however, not all enteroendocrine cells have such a quality, and their identification remains elusive unless special methods are used. Silver (argyrophilic, argentaffinic) preparations or immunocytochemical techniques are useful for visualization in these cases. The disadvantage of silver stains is that no information can be gleaned with respect to endocrine cell type or hormonal composition, and some cells do not reduce silver preparations (10). Immunocytochemistry is, therefore, the best method for definitive evaluation. Antibody preparations to chromogranin and neuron-specific enolase have been shown to be useful nonspecific markers for endocrine cells (36) (Figure 24.11). Precise identification of endocrine chemical content requires specific monoclonal or polyclonal antibody preparations. Although most often used in experimental settings, specific hormonal content immunostaining also may have diagnostic value in studying neuroendocrine tumors (37). Electron microscopy
P.609
also can be used to identify cytoplasmic neurosecretory granules.
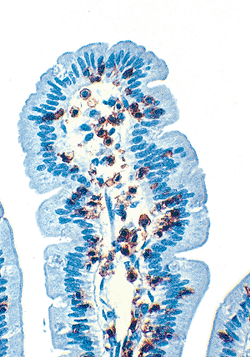 |
Figure 24.9 Small intestinal mucosa immunostained for UCHL1 (CD45 RO), a pan T-lymphocyte marker. The positive red-brown reaction highlights both the intraepithelial and lamina propria T cells. Although CD8-positive T-suppressor/cytotoxic lymphocytes predominate within the epithelium, lamina propria T cells usually mark for CD4, the T-helper/inducer subset. |
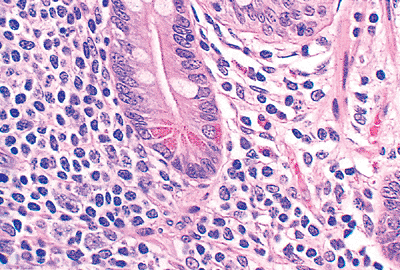 |
Figure 24.10 A single crypt surrounded by normal cellular lamina propria with abundant migratory cells. Both absorptive columnar and goblet cells are seen lining the crypt. In addition, an endocrine cell (infranuclear eosinophilic granules) and several Paneth's cells (supranuclear granules) are clearly evident. |
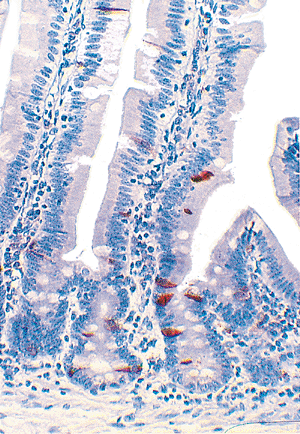 |
Figure 24.11 Immunostain for chromogranin shows numerous endocrine cells within the crypts and several scattered along the villous surface. |
At least 16 distinct types of endocrine cell have been described along the gut (38) and have a characteristic regional distribution and composition. Individual cells containing cholecystokinin, secretin, gastric inhibitory polypeptide, and motilin populate more proximal segments of the small bowel, whereas enteroglucagon-, substance P , and neurotensin-storing cells are seen in greater frequency in the ileum (38,39). Serotonin- and somatostatin-containing cells are not as regionalized and are found throughout the gastrointestinal tract (38). Some of these endocrine cells are known to play key roles in daily gastrointestinal activity. For example, secretin and cholecystokinin are released in response to various foodstuffs and modulate pancreatic secretion and gallbladder function, respectively (39). Most other gut endocrine cells are still of uncertain or unknown physiologic significance (11,39).
The Paneth's cells, normally found only in the crypt, comprise most of the base of individual crypts throughout the entire small intestine (40). They also are encountered to a lesser degree in the appendix, cecum, and ascending colon (41). Paneth's cells have a pyramidal shape with their apices pointing toward the lumen. Their cytoplasm contains characteristic supranuclear, intensely eosinophilic granules that are easily visualized in H&E stained sections (Figure 24.10). Interestingly, fixatives containing picric acid (e.g., Hollande's, Bouin's) mask the eosinophilic staining of these granules, often disclosing only unstained cytoplasmic vacuoles (40) (Figure 24.12). Their round nuclei often contain a prominent nucleolus (20,29). The exact function of Paneth's cells is still uncertain, but they are known to contain lysozyme, defensins (Figure 24.13), and immunoglobulin, and they are capable of phagocytic activity.
P.610
These features suggest that they play a role in regulating the intestinal microbial flora (41,42,43).
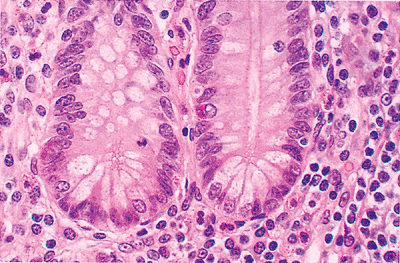 |
Figure 24.12 Although Hollande's and other picric acid containing fixatives are superior preservers of cytomorphologic detail, the characteristic supranuclear eosinophilic granules of Paneth's cells are not as easily visualized when compared with the appearance in 4% formaldehyde solution (see Figure 24.10). Clear vacuoles replace the distinct eosinophilic granules within the Paneth's cells (along the crypt bases). Note that the single endocrine cell in this crypt maintains its infranuclear granular staining quality. |
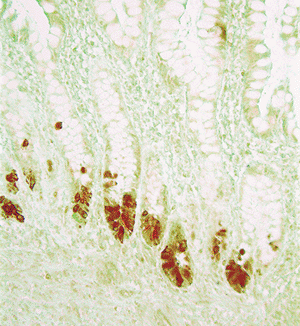 |
Figure 24.13 Immunostain for defensin HD5 highlights Paneth's cells at the base of the crypts. |
The crypt epithelium also contains intraepithelial lymphocytes that are predominantly T-suppressor/cytotoxic (CD8-positive) cells (44). Other inflammatory cell types, such as the neutrophil or the plasma cell, are not normally present within either the crypt or villous epithelial compartments; their presence would indicate a pathologic state (31,45).
Immunostaining Patterns of the Epithelium.
Carcinoembryonic antigen (CEA) is present on the apical surfaces of cells covering the villi and lining the crypts, and it has been shown to localize to the glycocalyx surface component (46). Additionally, the mucin droplets of goblet cells contain an abundance of CEA and consequently mark intensely with polyclonal anti-CEA. However, no intracytoplasmic immunostaining for CEA is evident in the normal small bowel (46).
Human leukocyte antigen (HLA)-DR like antigens have been shown to be present in a scattered, focal distribution on the apices of small intestinal columnar-shaped cells (47). Immunostaining with anti HLA-DR discloses a diminishing intensity of reactivity from the villous surfaces to the crypt bases. Immune-related cells such as lymphocytes (mostly B cells) and macrophages along with the walls of capillaries in the lamina propria also show immunoreactivity for HLA-DR (47).
Lamina Propria
The lamina propria, the intermediate layer of the mucosa, functions both structurally and immunologically. It rests upon the muscularis mucosae, surrounds the crypts, and extends upward as the cores of the intestinal villi. The crypt epithelium and villous epithelium rest upon the lamina propria and are separated from it by a distinct basement membrane recognized as a slender eosinophilic, PAS-positive band at their interface (Figures 24.4,24.5,24.6). This subepithelial basement membrane is a continuous structure composed of an ultrastructurally apparent basal lamina and a deeper network of collagenous/reticular fibers and ground substance (17). Interweaving collagen bundles and other connective tissue fibers, fibroblasts, mature fibrocytes, and smooth muscle cells comprise the framework of the lamina propria (45), whereas blood capillaries, lymphatics, and nerves course through this layer on their various routes to and from all portions of the bowel wall.
The most conspicuous feature of the lamina propria is its abundant immunocompetent and migratory cell component (Figures 24.10, 24.12). Five types of immunocompetent or inflammatory cells are normally encountered in the lamina propria: plasma cells, lymphocytes, eosinophils, histiocytes, and mast cells. Neutrophils are not usually encountered in the lamina propria, or for that matter anywhere in the small bowel wall, with the exception of those confined to vascular lumina.
Plasma cells are the most abundant cellular lamina propria constituent; most contain cytoplasmic immunoglobulin (Ig)A but some contain IgM (Figure 24.14). In contrast to extraintestinal sites (e.g., peripheral lymph nodes) (30,48), IgG-secreting plasma cells are scant. Lymphocytes, both B and T cells, are also common. The predominant subset of T cells within the lamina propria is the T-lymphocytes with the helper/inducer immunophenotype (CD4 positive) (27,48,49) (Figure 24.9). Lymphocytes are found throughout the lamina propria but often form more dense infiltrates just above the muscularis mucosae (45). Moreover, lymphoid aggregates and nodules, many with germinal centers, are scattered along the small bowel and are found in increasing concentration distally. These lymphoid aggregates are based in the lamina propria, but they often extend to some degree into the underlying submucosa.
Histiocytes or macrophages are also seen in the lamina propria but in fewer numbers than lymphocytes or plasma cells. Most are located along the superiormost aspect of the lamina propria near the tips of the villi (28). They function in T-cell regulation as antigen presenters and phagocytes (30).
P.611
Exaggerated expression of this latter function can be seen pathologically in disseminated Mycobacterium avium-intracellulare complex infection and in Whipple's disease, where the lamina propria becomes filled with macrophages engorged with microorganisms (30,50,51,52).
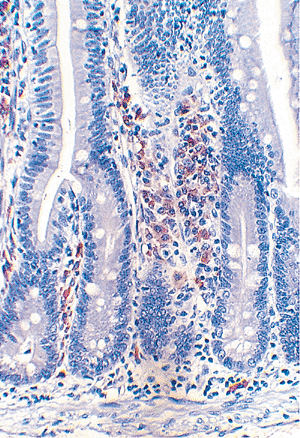 |
Figure 24.14 Numerous IgA-containing plasma cells (red-brown cytoplasmic staining) within the lamina propria of normal small intestine, immunostained for -heavy chain. Note that, in contrast to T cells (see Figure 24.9), the plasma cells localize to the lamina propria and are not normally found in the epithelium. |
The only granulocytes normally found in the lamina propria are eosinophils and mast cells. Eosinophilic leukocytes usually are conspicuous, but their role in the normal bowel is uncertain (53). The numbers of eosinophils in the lamina propria increase under various conditions, including those comprising the eosinophilic gastroenteropathies (54,55,56,57,58). Mast cells are relatively abundant in the small intestinal lamina propria, as compared with other body sites (30), but in absolute numbers appear to decrease with distal progression along the small bowel (59). Their function in the gastrointestinal tract in normal and disease states is unknown. Increased numbers of mast cells can be found in inflammatory bowel disease and in the specimens of some individuals with eosinophilic gastroenteropathy (57,60). According to some investigators, a specimen disclosing more than eight mast cells per high-magnification field should suggest the diagnosis of systemic mast cell disease (61). Mast cells can be seen on H&E stained sections, but they are more easily visualized with special techniques such as toluidine blue, sulfated Alcian blue, or Giemsa stains. Rarely, subepithelial (lamina propria) endocrine cells may be found in the small bowel; however, these are much more prominent in the vermiform appendix (62). Occasionally, in apparently healthy individuals and in certain disease states (e.g., Crohn's disease), ganglion cells are found in the lamina propria of the small bowel. These could potentially be confused with cytomegalovirus infection induced cellular changes.
Muscularis Mucosae
The muscularis mucosae, which is the outermost layer or limit of the mucosa, is a slender band of tissue composed of elastic fibers and smooth muscle arranged in an outer longitudinal and inner circular layer (Figure 24.15). However, these layers are usually not well delineated on routine light microscopy. Tufts of smooth muscle radiate from the muscularis mucosae into the lamina propria and extend into the villi. The muscularis mucosae provides an important structural foundation for the mucosa, and its absence in some biopsy specimens can cause a loss of villous orientation, an artifact that may interfere with optimal evaluation (53).
Submucosa
Between the muscularis mucosae and muscularis externa is the submucosa, a loose, paucicellular layer composed of a regular, honeycomb-like arrangement (at the ultrastructural level) of collagenous and elastic fibers and related fibroblasts. The submucosa also may contain scattered, rather inconspicuous migratory cells (e.g., histiocytes, lymphoid and plasma cells, and mast cells) and adipose tissue (63). Its histologic appearance and principal role in maintaining the structural integrity of the small bowel are similar throughout the gastrointestinal tract (63). The submucosa is a major focus of vascular routing and related distribution of regional blood and lymphatic flow. Relatively large caliber arterioles, venules, and lymphatic vessels form extensive individual plexuses and networks within this layer (5,19) (Figure 24.16). From this vascular center, numerous penetrating capillary vessels supply and drain most of the mucosa and muscularis externa (5,19). Lymphatic vessels may be distinguished from blood vessels by the thinner wall of the former and the lack of luminal erythrocytes. However, certain immunohistologic patterns and electron microscopic characteristics are more helpful for definitive identification (18,64,65). Specifically, the endothelial cells of blood capillaries immunostain for PAL-E and factor VIII related antigen, whereas lymphatic capillary endothelia typically lack these antigenic sites and remain unstained with such antibody preparations (64,65). In addition, blood capillaries as seen by ultrastructural analysis have a continuous basal lamina, endothelial fenestrations, and ensheathing pericytes. However, lymphatic capillaries have a discontinuous basal
P.612
lamina and lack both fenestrations and surrounding pericytes (17,18). Although small lymphatic vessels are a conspicuous submucosal component, prominent dilated lymphatic structures in this layer, as well as in the mucosa, can be seen in pathologic states such as intestinal lymphangiectasia or Crohn's disease (31,66).
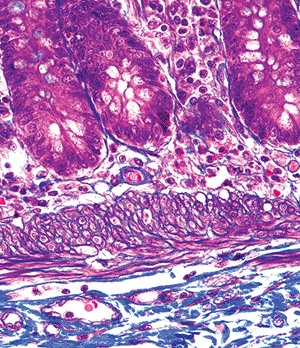 |
Figure 24.15 High-magnification photomicrograph disclosing inner circular and outer longitudinal smooth muscle bands of the muscularis mucosae; this layering is often inconspicuous on hematoxylin and eosin preparations, where the muscularis mucosae appears as a thin eosinophilic strip between the lamina propria and underlying submucosa (Masson's trichrome). |
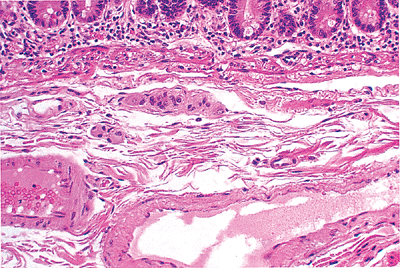 |
Figure 24.16 Normal submucosa separated from overlying mucosa by the eosinophilic-staining muscularis mucosae. The submucosa is paucicellular, disclosing fibrocollagenous tissue and a prominent vascular component. Note the ganglion cells of Meissner's plexus just beneath the muscularis mucosae. |
Neural structures are also prominent in the submucosa. The submucosal Meissner's plexus forms one of the two major integrative centers of the enteric nervous system. It consists of a network of ganglia that interconnect through neural processes (67). The ganglia contain compact aggregates of neurons (ganglion cells) routinely identified on H&E preparations by their characteristic large oval shape, abundant pink cytoplasm, vesicular nucleus, and single prominent, often eosinophilic nucleolus (Figure 24.16). Abundant S-100 positive Schwann cells, glial-like cells, and neural processes are also present in Meissner's plexus. The entire plexus, including the ganglia, contains no connective tissue elements or vascular structures in the normal state (67,68,69,70). The plexus is also normally devoid of inflammatory cells; therefore, if these are seen, an injury pattern specific to the neural plexus (such as an inflammatory neuropathy) should be considered, as long as primary inflammatory bowel disease can be excluded (69). Neural interconnections exist between Meissner's plexus and the myenteric plexus of Auerbach (discussed below), as well as with extrinsic (autonomic) neural processes.
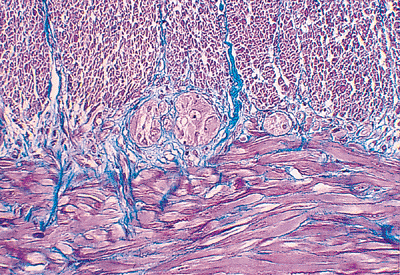 |
Figure 24.17 Masson's trichrome stain clearly delineates the inner circular (above) from the outer longitudinal (below) smooth muscle bands of the muscularis externa. The prominent muscular component (red) is partitioned into bundles of varying size by delicate collagenous fibers (blue). Note the ganglia of the myenteric plexus of Auerbach, characteristically located between the two muscle bands. Fibrous tissue is minimal within the muscularis externa and is also not normally part of the plexus. |
Muscularis Externa
The muscularis externa (or muscularis propria) is the thick outer smooth muscle layer that surrounds the submucosa. It is covered externally by subserosal connective tissue and, in most places, by a serosa. Its two distinct muscular layers, oriented perpendicular to each other, are arranged as an outer longitudinally running muscle fiber layer and an inner circular muscle band (Figure 24.17). Blood vessels, lymphatics, and nerves course through the muscularis externa and slender collagenous septa surround groups of smooth muscle cells, creating characteristic bundles and packets of muscle (Figure 24.17). However, fibrous tissue in this layer is minimal in the normal small bowel (69) so even slight fibrous alterations or collagen deposition may be significant. Moreover, the fact that only a few disease entities (including ischemia, irradiation, familial visceral
P.613
myopathy, scleroderma, and mycobacterial infection) are associated with fibrosis of the muscularis propria aids in narrowing a broad differential diagnosis (31).
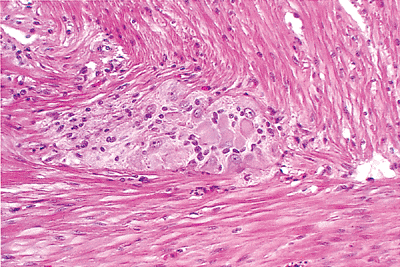 |
Figure 24.18 A single ganglion of the myenteric plexus of Auerbach located at the interface of the inner (above) and outer (below) smooth muscle layers of the muscularis externa. Ganglion cells (neuronal cell bodies) are evident and characterized by a polygonal shape, abundant pink cytoplasm, and an eccentric nucleus; spindled neural projections and Schwann cells are also intermixed. |
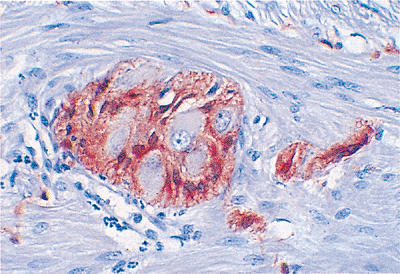 |
Figure 24.19 This S-100 immunostain highlights the otherwise inconspicuous spindled Schwann cell component of the ganglion. It also marks the Schwann cells accompanying the neural projections that interconnect these ganglia to one another within the plexus system. Note that the ganglion cells show no such immunoreactivity. |
The myenteric plexus of Auerbach, the other major neural plexus of the enteric nervous system, lies between the outer longitudinal and inner circular muscle layers (Figures 24.17,24.18,24.19). Auerbach's plexus is similar in composition to the submucosal plexus, although it typically has larger ganglia, a greater number of neurons, and a more compact plexus network (68,69). As a consequence of these features, it is best to evaluate the myenteric plexus for specific disease processes involving the enteric nervous system, such as the various visceral neuropathies. Because routine processing allows only a small portion of the plexus to be visualized and because many of these conditions cause no detectable changes on routine H&E-stained sections, special preparations of thicker, larger, and silver-stained sections cut en face are currently necessary to diagnose many of these disorders (69). Finally, although of lesser importance, a deep muscular, subserous plexus and several mucosal plexuses are also present within the small bowel (68). Interstitial cells of Cajal (ICC) are located in relation to the myenteric plexus of Auerbach and in septa between circular muscle lamellae (Figure 24.20). These pacemaker cells are felt to play a role in intestinal motility (71).
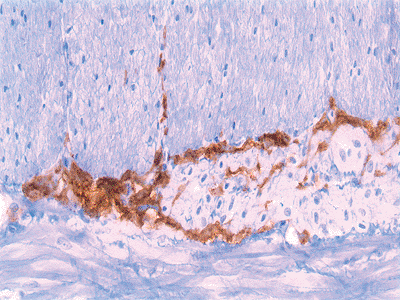 |
Figure 24.20 The CD117 (c-kit) immunostain shows interstitial cells of Cajal surrounding the myenteric plexus of Auerbach. |
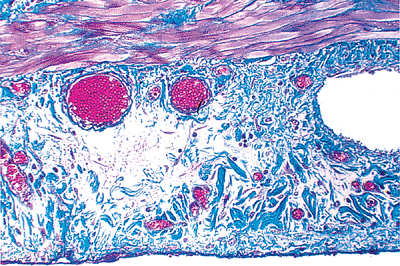 |
Figure 24.21 The subserosal region contains a delicate fibrocollagenous network, blood vessels, lymphatics, and nerves. The serosa consists of a thin fibrous layer (blue and bottom) covered by a single layer of mesothelial cells; however, the mesothelium is often denuded in surgical specimens. A portion of the outer layer of the muscularis externa is also present in this field (top). (Masson's trichrome.) |
Serosa and Subserosal Region
The serosa is the covering that envelops most of the external surface of the small bowel. Its outermost layer consists of a single row of cuboidal mesothelial cells, under which lies a thin band of loose connective tissue. A subserosal zone of connective tissue lying between this mesothelial covering and the muscularis externa also contains ramifying branches of blood vessels, lymphatics, and nerves (Figure 24.21).
Distinctive Regional Characteristics of the Small Bowel
Duodenum
The duodenum exhibits several distinctive histologic features, many related to its proximal location in direct continuity with the pylorus. The gastroduodenal junction,
P.614
although well-delineated grossly, is poorly demarcated histologically (72) (Figures 24.22,24.23). A gradual transition in epithelial types occurs, with three distinct subtypes in the duodenum (73): (a) an antral-type mucosal epithelium that is identical to the pyloric mucosa; (b) a usual small intestinal type (jejunal type) characterized by villi covered by absorptive cells and interspersed goblet cells; and (c) a transitional type (Figure 24.23), in which the same villus is covered by epithelium having features of both antral-type and usual small intestinal-type epithelia. In the region of the gastroduodenal junction, irregular undulating slips of antral-type mucosa extend about 1 to 2 mm into the anatomic duodenum, which then abuts a 2- to 3-mm segment of transitional-type epithelium (73). Distal to this, only the usual small intestinal-type mucosa is found (73). The transitional type epithelium occurring in more distal aspects of the duodenum and in the rest of the small intestine is termed gastric metaplasia (31).
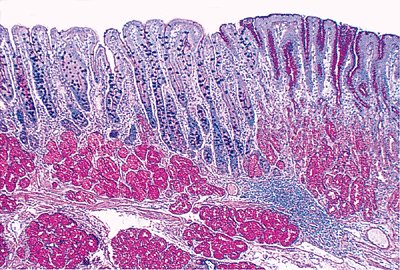 |
Figure 24.22 Gastroduodenal junction. Note the transition from PAS-positive (red) gastric foveolar epithelium and underlying pyloric glands (right) to a villous mucosal architecture of the duodenum (left) lined predominantly by Alcian blue/PAS-positive (blue-purple) goblet cells and absorptive cells. Note that both pyloric (right) and Brunner's glands (left) are composed predominantly of cells containing only neutral, PAS-positive mucin. Brunner's glands, however, are predominantly submucosal in location, while pyloric glands are an intramucosal structure. (Alcian blue/PAS.) |
Although the duodenal mucosa may demonstrate long villi with a villous-to-crypt length ratio on the order of 3:1 to 5:1, more commonly, particularly in the first portion (the duodenal cap or bulb), the villi are shorter and broader with occasional branching extensions (74) (Figure 24.24). They often have a leaflike shape with few fingerlike forms when viewed under a scanning electron or dissecting microscope (20,74,75) (Figure 24.25). Also, the number of mononuclear cells within the lamina propria is increased in the duodenum when compared with the rest of the proximal small intestine (74,75). This varied constellation of findings is considered normal and is probably a consequence of the effect of acidic gastric contents on this most proximal intestinal site (40,75).
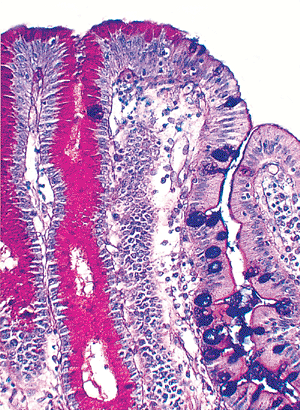 |
Figure 24.23 Several villi within the confines of the gastroduodenal junction disclosing both usual small intestinal-type epithelium and antral-type, PAS-positive, foveolar epithelium. This transitional-type epithelium is a characteristic hybrid found in this region. At more distal small intestinal sites, this transitional type epithelium is termed gastric metaplasia. (Alcian blue/PAS.) |
The submucosa of the gastrointestinal tract lacks glands except at two sites: the esophagus and the duodenum. The submucosal Brunner's glands are the type localized to the duodenum. Indeed, these glands are typically used by the pathologist to identify histologically a segment of small
P.615
intestine as duodenum. Brunner's glands, which begin just distal to the gastroduodenal junction, are most concentrated in this region and gradually decrease in quantity along the duodenum (76). Beyond the entrance of the ampulla of Vater, only scattered groups can be found. In rare instances, Brunner's glands extend beyond the duodenojejunal flexure for a short distance (77,78,79).
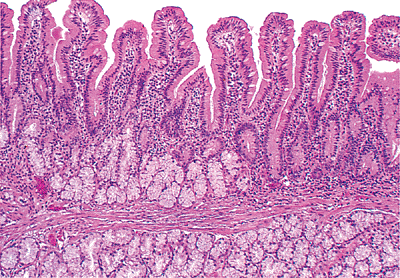 |
Figure 24.24 Short, slightly broader villi predominate in the duodenum. The underlying submucosal Brunner's glands are a distinctive feature of this portion of small bowel. Note that a fair portion of Brunner's glands normally occurs above the muscularis mucosae. |
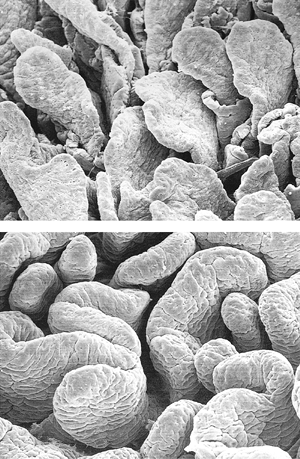 |
Figure 24.25 Two views of duodenal mucosa using scanning electron microscopy: leaflike (A) and ridged-shaped (B) villi predominate in these normal duodenal specimens. |
Brunner's glands are lobular collections of tubuloalveolar glands predominantly located within the submucosa; however, they often extend through the muscularis mucosae into the deep portions of mucosa beneath the crypts of Lieberk hn (Figures 24.24, 24.26). On average, about one third of the gland population resides within the mucosa (77). Brunner's glands are lined by cuboidal-to-columnar cells with pale, uniform cytoplasm and an oval, basally situated nucleus. Their cytoplasm contains neutral mucins that are PAS positive and diastase resistant (Figure 24.22). Occasionally, mucous cells with apically concentrated mucin and perinuclear vacuolization or clearing are seen. Although opinions vary, these changes are thought to represent the secretory phase of the gland (i.e., recently fed state) (80,81). The glands empty by way of ducts lined by a similar epithelium, which are often seen passing through slips of muscularis mucosae (Figure 24.26). These ducts drain into the crypts at varying levels (79). Brunner's glands and their ducts can be distinguished from surrounding crypts by the absence of goblet cells and by their diffuse cytoplasmic PAS positivity (79).
Although most of the lining epithelial cells of the Brunner's glands are of the mucous type, scattered
P.616
endocrine cells are present as well. Many can be detected on routine H&E-stained sections because of their basal eosinophilic granulated cytoplasm (82). By using immunohistologic methods, some have been shown to contain somatostatin, gastrin, and peptide YY (83). However, the ducts that drain Brunner's glands are devoid of endocrine cells (83).
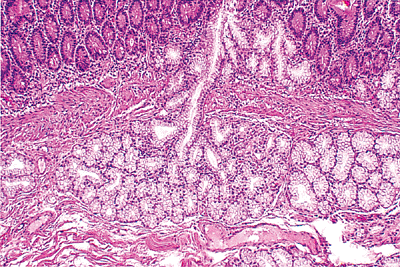 |
Figure 24.26 Submucosal Brunner's gland lobule with draining duct extending through muscularis mucosae. Note the stark contrast between the crypt epithelium and that of the Brunner's glands and their ducts. |
Peptidergic neural fibers, predominantly those with immunoreactivity for vasoactive intestinal peptide and substance P, course within and between individual Brunner's glands. These neuroendocrine substances are probably important in local regulation of acinar secretion, although this function has been verified only for vasoactive intestinal peptide (83). The function of Brunner's glands has not been fully elucidated, but their mucus is felt to be of prime importance for protection of the duodenal mucosa from the potentially damaging effects of the delivered acidic gastric contents (78).
Hyperplasia of Brunner's glands exists in three forms: (a) diffuse glandular proliferation, imparting a coarse nodularity to most of the duodenum; (b) isolated discrete nodules in the proximal duodenum; and (c) a solitary nodule, often designated as an adenoma of Brunner's glands (77,84,85). All three types are typically composed of an increased quantity of normal-appearing Brunner's glands, accompanied by variable proportions of smooth muscle (Figure 24.27). The distinction between adenoma and hyperplasia is arbitrary, and no substantial evidence exists to suggest that any of these proliferations are truly neoplastic (84). Moreover, carcinoma arising from a population of Brunner's glands has yet to be convincingly documented (31). Nodules or polypoid structures composed of collections of these submucosal glands in the duodenum are probably best termed Brunner's gland nodules (31).
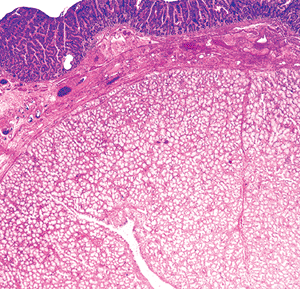 |
Figure 24.27 Brunner's gland nodule disclosing abundant normal-appearing submucosal glands of Brunner intermixed with smooth muscle, underlying an unremarkable duodenal mucosal villous surface. |
Pseudomelanosis duodeni, or brown-black pigment, located primarily within lamina propria macrophages, rarely may be observed in the proximal duodenum (86) (Figure 24.28). Lipomelanin, ceroid, iron, sulfide, and hemosiderin have been identified in these deposits. Most reported patients were hypertensive and also suffered from upper gastrointestinal bleeding, chronic renal failure, or diabetes mellitus (86).
Jejunum
The jejunum is the least distinctive segment of the small bowel; and, as such, its histologic features are most similar to those described for the small bowel in general. However, a characteristic feature is the prominent development of the plicae circulares, or permanent circular folds, also termed valves of Kerckring and valvulae conniventes (Figure 24.1). These folds are tallest and most numerous (i.e., closely spaced) in this portion of the small bowel (20). Histologically, the jejunal villi are tall with a villous-to-crypt ratio on the order of 3:1 to 5:1. The vast majority of jejunal villi are slender and fingerlike (Figures 24.2,24.3), in contrast to the slightly shorter villi of the ileum and to the leaflike, occasionally branched and blunted villi of the proximal duodenum (20,40). These morphologic transitions are gradual, particularly in the mobile small intestine, where the separation between jejunum and ileum is arbitrarily defined.
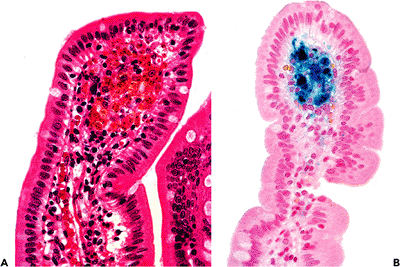 |
Figure 24.28 A. Macrophages containing granular brown-black pigment within the lamina propria of the duodenum, a characteristic of pseudomelanosis duodeni. B. Prussian blue stain disclosing the prominent iron content of the pigment. |
P.617
Ileum
The ileum has a number of distinctive features, including its unique junction with the large intestine. The ileum protrudes approximately 2 to 3 cm into the large intestine at the junction of the cecum and ascending colon. This nipplelike extension of the terminal ileum is encircled by large bowel mucosa and has been likened to the relationship of the uterine cervix with the vagina (87). A muscular sphincter at this site, along with external ligamentous support, is responsible for modifying its function in order to prevent reflux and to allow forward passage of ileal contents (87,88). Histologically, the mucosal transition demonstrates a gradual loss of villi occurring at variable lengths along the short intracecal ileal segment; the ileal mucosa blends rather imperceptively with the mucosa of the large bowel (Figure 24.29). The ileocecal region normally can contain abundant fat within its submucosa, diffusely distributed and proportional to adipose content in the rest of the abdominal cavity (89) (Figure 24.29). In fact, on rare occasion, a distinct mass of fat is evident. This benign entity, so-called lipohyperplasia of the ileocecal region, reportedly can cause variable symptoms, including abdominal pain and lower gastrointestinal bleeding (89).
The distinctive mucosal characteristics of the ileum, when compared with both jejunum and duodenum, include shorter and fewer plicae circulares and an increased proportion of goblet cells within the epithelium (Figure 24.30). The villi are typically shorter than at more proximal sites and have a predominantly fingerlike shape (20). These features become gradually more apparent along the length of the small intestine and are most evident in the distal ileum. With distal progression, lymphoid nodules that can be found anywhere in the small intestine gradually increase in quantity (20). In addition, specialized clusters of lymphoid aggregates, or Peyer's patches, are most prominent in the ileum.
Peyer's patches are located along the antimesenteric border of the small bowel. They consist of varying numbers of lymphoid follicles, ranging from 5 to over 900, and have been shown to be present during fetal life (90). Until puberty, they increase in size and number and subsequently regress steadily thereafter; nonetheless, Peyer's patches are invariably present even into extreme old age (90). In children, these patches can be grossly visualized near the
P.618
ileocecal junction (91); although on the whole, they cannot be seen with the naked eye (92). Moreover, hyperplastic Peyer's patches (or focal lymphoid hyperplasia) may be found in the terminal ileum during childhood and have been linked to more than one-third of the cases of idiopathic intussusception occurring in the ileocecal region in this age group (93,94,95).
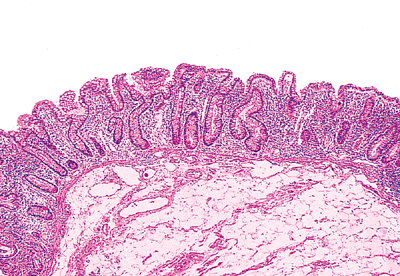 |
Figure 24.29 Transition from villous mucosal surface of ileum (left) to flat mucosa of large intestine (right) at the ileocecal junction. Note the prominent submucosal adipose tissue characteristic of this region. |
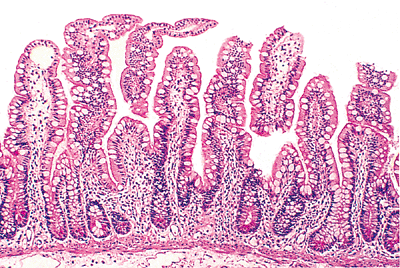 |
Figure 24.30 Characteristic ileal mucosa with slender relatively short villi (compare with jejunal villi in Figure 24.3) lined by abundant goblet cells with a lesser number of absorptive columnar cells. |
Peyer's patches are basically specialized groups of lymphoid follicles that occupy the mucosa and a variable portion of the submucosa. Structurally, four distinct compartments exist: follicle, dome, follicle-associated epithelium, and interfollicular regions (30,96) (Figure 24.31). Most lymphoid follicles within a Peyer's patch contain a germinal center that is populated by numerous surface IgA-positive B cells, with occasional CD4-positive T cells and macrophages (30,92) (Figures 24.31,24.32). However, the surrounding mantle zone contains a population of small B cells (predominantly surface IgD- and IgM-positive). The dome is the area between the follicle and the overlying surface epithelium; this region contains a heterogeneous population of cells, including B cells of all immunoglobulin isotypes (except IgD), macrophages, and plasma cells (92). The specialized or follicle-associated epithelium overlying lymphoid aggregates is distinct from surrounding villous epithelial surfaces. It characteristically has fewer goblet cells and contains membranous cells, or M cells, interspersed among the usual columnar absorptive cells (30,97) (Figure 24.32). The M cell is a specialized columnar epithelial cell that transports luminal antigens to adjacent extracellular spaces, thus allowing access to immunocompetent cells. These cells play a key role in mucosally based immunity, antigen tolerance, and probably in certain immunopathologic disease states (97). Morphologically, M cells have an attenuated brush border with diminished alkaline phosphatase staining intensity (96,98), a thin strip of apical cytoplasm, and a basally situated round nucleus. The M cell's cytoplasm is often deformed by several lymphocytes (97). However, definitive M-cell characterization rests upon the identification of specific ultrastructural features (96,97). The follicle-associated epithelium also has a distinctive migratory cell composition because the intraepithelial lymphocyte population is more abundant and has a greater proportion of CD4-positive (T-helper/inducer) cells than does the usual villous epithelium (approximately 40% versus 6%) (96). Last, the fourth component of the Peyer's patch is called the interfollicular region. As in the lymph node, it is predominantly a T-cell zone with CD4-positive T-lymphocytes outnumbering CD8-positive lymphocytes by a 7:1 margin (92).
Plasma cells are a scant component of Peyer's patches, in contrast to the surrounding lamina propria, but they may frequently be found in the dome compartment. Peyer's patches are believed to be important in the generation of the mucosal immune response, in part by supplying the lamina propria with immunocompetent surface IgA-positive B cells
P.619
that become functional secretory plasma cells (30,91,92,97).
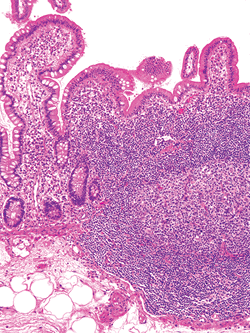 |
Figure 24.31 Confluent lamina propria lymphoid aggregates of a Peyer's patch. This organized lymphoid tissue typically extends into the underlying submucosa. The four components of the Peyer's patch are seen and include lymphoid nodules with germinal centers, an overlying flattened follicle-associated epithelium, and an intervening pale-staining dome region. The abundant lymphoid population between the follicles is the T-cell rich interfollicular zone. |
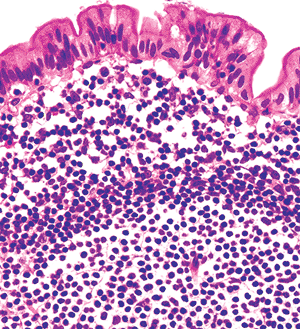 |
Figure 24.32 High magnification of surface epithelium above a lymphoid nodule within a Peyer's patch. The polymorphous germinal center (below) is surrounded by monotonous, small round lymphocytes that comprise the nodule's mantle zone. Above this lies the dome region with lymphocytes, plasma cells, and macrophages. The follicle-associated epithelium characteristically has few, if any, goblet cells; ultrastructurally, most of these cells would be identified as M cells. |
Irregularly distributed, granular brown-black pigment can commonly be found in the deep portions of Peyer's patches in adults (99) (Figure 24.33). Although its origin is controversial, atmospheric or dietary sources are most probable (99,100). Accumulating principally within macrophages, this pigment has been shown by x-ray spectroscopy to contain a distinct mineral composition that includes silicates, aluminum, and titanium (99,100). The pigment is inert and has no known clinicopathologic significance.
A final distinctive feature often seen in the ileum is Meckel's diverticulum; it is the most common intestinal congenital anomaly and is found in 1 to 2% of the general population (31). Meckel's diverticulum is an antimesenteric outpouching of the terminal ileum usually located approximately 20 cm from the ileocecal junction. It represents the persistence of the omphalomesenteric duct. Although Meckel's diverticulum is usually an incidental finding, it can cause lower gastrointestinal bleeding or small bowel obstruction (101,102). Histologically, small intestinal mucosa alone lines the diverticulum in about 50 to 70% of cases. Ectopic gastric or pancreatic tissues are found in the remainder, typically encountered at the distal most aspect (102).
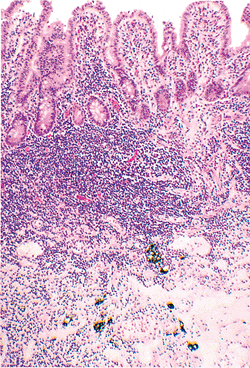 |
Figure 24.33 Dense brown-black granular pigment within the depths of a Peyer's patch of ileum. The pigment is typically confined to macrophages. |
Special Considerations
Geographic, Age-Related, and Dietary Factors
Because geographic and local environmental factors can affect small bowel morphology, historical data about the residence or recent travel of an individual are essential to evaluate histologic material accurately. Specimens from individuals residing in, or visiting at length, certain less developed tropical locations such as Africa, southern India, and Thailand show a distinctly different villous appearance from those of individuals who live in temperate climate zones (45,103,104,105,106). The morphologic alterations seen in individuals from such tropical areas include leaflike villi predominating over fingerlike forms in jejunal segments examined via biopsy and an increased number of lamina propria mononuclear cells (107). This difference in the villous population is reflected histologically as stubby villi with a pyramidal shape (i.e., a broader base than apex) and occasional branched and fused villous tips (45,103,104). Interestingly, although the villi are shorter, the villous-to-crypt length ratio usually remains constant in all geographic settings (45,104). Such alterations should probably be considered a normal variant because these individuals are typically asymptomatic and otherwise healthy (103).
P.620
The cause of these morphologic changes is uncertain, but environmental factors, particularly regional enteric flora, presumably play a role (103). Similar mucosal changes are seen in normal individuals in temperate environments, but only in proximal portions of the duodenum. Therefore, these particular mucosal alterations must be analyzed in the context of both the patient's residence and the site within the small intestine in order to prevent misinterpretation as a pathologic change, such as tropical sprue.
Aging also modifies small bowel mucosal architecture. Although the literature on humans is limited, it has been shown that specimens from elderly individuals generally have shorter and broader villi than those from younger individuals (108). Moreover, lower animal and human fetuses have been documented as having fingerlike villi exclusively, suggesting that exposure to the environment or aging itself modifies villous architecture (108,109). However, the functional significance of these changes is uncertain (108).
Diet alters villous architecture in laboratory animals. A diet high in fiber results in broad and fused villi, whereas a fiber-free diet seems to prevent the formation of leaflike forms (17). If this finding is valid in humans, it may be one factor related to the presence of stubby, leaflike villi seen in patients in less developed countries where high-fiber diets are common.
Metaplastic and Heterotopic Tissues
Gastric type mucosa is not an unusual finding in the small intestine. A distinction can be made between metaplasia, an acquired alteration, and heterotopia, thought to be congenital in origin. Gastric metaplasia characteristically consists solely of antral-type, PAS-positive, foveolar columnar cells lying along the surface epithelium (Figure 24.23). This change is focal and often in direct continuity with usual columnar absorptive epithelium on the same villus (75). Gastric metaplasia may be encountered in more than 60% of healthy asymptomatic individuals in the duodenal bulb, where it can be regarded as within normal limits (75). More distally, however, it is less commonly seen in the asymptomatic person but rather frequently is associated with duodenitis or mucosal ulceration (75,110). Scattered chief and parietal cells without any organized arrangement are also associated with this type of metaplasia or with reparative processes (110).
In contrast, gastric heterotopia is usually a grossly evident mucosal polyp that contains all cellular elements encountered in the normal gastric fundic mucosa. Characteristically, mucous foveolar epithelium overlies an organized arrangement of glands lined by chief and parietal cells; this is typically well demarcated from the surrounding usual intestinal villous epithelium. Gastric heterotopia is also fairly common, being reported in up to 2% of the population (111); it may be found anywhere along the gastrointestinal tract (110). Gastric heterotopia is a well-defined entity in the proximal duodenum and usually presents as a mucosal nodule on the anterior wall. Although they are usually of no clinical significance, larger ones may cause obstructive symptoms (111). In contrast, gastric heterotopia distal to the ligament of Treitz is usually symptomatic and often causes intussusception (112). This relationship to clinical symptoms may derive from patient selection bias because asymptomatic gastric heterotopias at a distal site would not be routinely detectable.
Heterotopic pancreas tissue also can be found anywhere along the small intestine but most commonly is found in the duodenum and jejunum (113,114). It can form submucosal, intramural, or serosal nodules and is composed of various admixtures of pancreatic acini, ducts, and islets of Langerhans (113,115) (Figure 24.34). Isolated ductal structures admixed with smooth muscle may be the predominant or exclusive component, and in these instances the alternative term adenomyoma has been used (31,114). Nodules of pancreatic tissue within the small intestine are usually asymptomatic, although larger lesions (greater than 1.5 cm) with prominent mucosal involvement may become clinically significant (114,115). A single case of a combined submucosal pancreatic heterotopia with overlying gastric type mucosa in the duodenal bulb recently was reported (116).
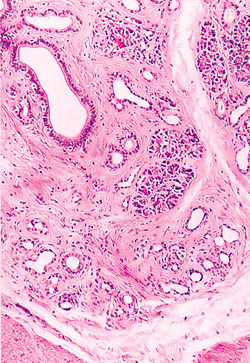 |
Figure 24.34 Heterotopic pancreas in the duodenum characterized in this instance by variably sized ducts, acini, and abundant smooth muscle. |
P.621
Lymphoid Proliferations
Lymphoid tissue is a prominent feature of the small bowel. The gut-associated lymphoid system in this region, as in the entire gastrointestinal tract, is compartmentalized into lymphocytes of the epithelium, lamina propria, isolated lymphoid aggregates, and Peyer's patches (117). The normal appearance and immunologic composition of these distinct lymphoid populations have been detailed earlier in the chapter. All these compartments participate at some level in mucosal immune response, but they also provide the milieu for various hyperplastic and neoplastic immunoproliferations, as well as for certain immunodeficiency states. Some of these disorders have histologic features that deviate only slightly from a normal appearance and from one another. Additionally, some are believed to be preneoplastic.
Lymphoid hyperplasias are divided into two broad categories: focal and diffuse forms (118). Focal lymphoid hyperplasia is a localized, well-circumscribed proliferation of benign lymphoid tissue characterized by a polymorphic infiltrate of lymphocytes within which numerous benign follicles with reactive germinal centers are dispersed. These proliferations often involve only the mucosa and submucosa, but they may extend through the entire wall (118). Focal lymphoid hyperplasia is predominantly found in the terminal ileum of children or adolescents who present either with ileocecal intussusception or with a clinical syndrome mimicking appendicitis (93,95,118).
Diffuse or nodular lymphoid hyperplasia is a distinct entity in which multiple nodules composed of aggregates of benign lymphoid follicles disfigure the mucosa and submucosa along extensive lengths of the small intestine, with or without colonic involvement (118,119,120). It is usually asymptomatic and incidentally encountered (118). However, a distinct clinicopathologic variant exists that is characterized clinically by associated, combined variable immunodeficiency and giardiasis-related diarrhea and histologically by a greatly diminished or absent plasma cell population in the nearby lamina propria (118). Nodular lymphoid hyperplasia, with or without combined variable immunodeficiency, has been associated with an increased risk for the development of various malignancies (e.g., malignant lymphoma, carcinoma) (118,119,120,121).
Differentiation of these benign lymphoid lesions from malignant lymphoma can be problematic. However, recognizing the diagnostic morphologic features of malignant lymphoma, such as the usual cytologic monotony of the lymphoid population, the lack of a reactive follicular architecture, and the presence of mucosal ulceration, should resolve most dilemmas (118,122,123,124). Additionally, because the majority of small bowel malignant lymphomas are of B-cell lineage (124), demonstration of light-chain restriction by immunohistologic techniques or flow cytometric analysis or detection of heavy- or light-chain immunoglobulin gene rearrangements (e.g., monoclonality) by molecular diagnostic methods can aid in certain cases (125,126,127); T-cell receptor gene rearrangement analysis may be necessary in a minority of cases.
Morphologic Changes Associated with Ileal Diversion and Continence-Restoring Procedures
With the increasing number of diversion and continence-restoring procedures being performed after total colectomy, it has become common to see biopsy and revision specimens after such operations. As a consequence, familiarity with the altered yet normal morphology within these ileal creations must be appreciated in order to evaluate them optimally. The expected mucosal changes include villous shortening and crypt lengthening (approximate 1:1 to 2:1 ratio), increased numbers of goblet cells and lymphoid follicles, and a denser mononuclear cell infiltrate within the lamina propria (31,128,129). These alterations are similar after either colectomy with conventional ileostomy or after ileoanal anastomosis with ileal reservoir formation (e.g., pouch) (129,130,131). However, ileostomy stomas in particular show additional changes of mucosal prolapse exemplified by fibromuscular obliteration of the lamina propria and superficial erosions and microhemorrhages (31). Additionally, goblet cell mucin alterations have been seen in nearly 50% of pouches examined, with conversion to predominantly sulfomucins (i.e., colonic epithelial mucin) (131). However, another group of investigators saw no change in goblet cell mucin from the typical small bowel acid sialomucins in ileal segments after either ileoanal anastomosis with pouch formation or conventional ileostomy (129). Nonetheless, all these changes should be interpreted as normal because more definitive and specific criteria need to be met to establish persistent, recurrent, or novel disease in these specimens.
Mucosal Biopsy Specimen Evaluation in Suspected Malabsorption
Specimen Procurement and Processing
The usefulness of small bowel mucosal biopsy is unquestioned (132), particularly in the evaluation of malabsorptive states. In the past, up to four biopsy samples were usually obtained from the area of the ligament of Treitz via a suction biopsy device attached to a long tube (133). In recent years, the standard upper endoscope has been used, and comparable specimens have been procured (76,134). Because this technique is performed under direct visualization, many
P.622
more biopsy specimens can be obtained. Regardless of the biopsy technique used, the most critical part of the procedure is proper orientation of the specimen. Ideally, specimens are immediately mounted mucosa side up on a solid substance such as filter paper or plastic mesh and then placed into Hollande's or other fixative. After processing, the histotechnologist embeds the tissue perpendicular to the mounting material. Alternatively, biopsy specimens may be placed unmounted into fixative immediately. The tissue can then be properly oriented after processing at the time of embedding. Because the specimen will naturally curl, some tangential sectioning can be expected. Proper specimen evaluation requires examination of optimally oriented intestinal villi obtained from the central region of the biopsy specimen. Although serial sectioning is advocated by some investigators (107), step-sectioning (three seven levels) is a reasonable alternative.
Our standard small bowel biopsy procedure consists of obtaining four to six endoscopic biopsy specimens. One can be used to make a touch preparation that is then fixed in alcohol and stained via the Giemsa technique. The other tissue samples are placed in fixative and routinely processed. Step-section slides are obtained: three are stained with H&E and one with alcizn blue/PAS. The PAS stain is a useful screen for Whipple's disease and Mycobacterium avium-intracellulare complex infection. Trichrome stain can be used to confirm collagen deposition seen in ischemia or collagenous sprue. In addition, the iron hematoxylin counterstain used in the trichrome technique makes it easier to identify giardiasis.
Specimen Interpretation and Common Artifacts
With appropriate specimen procurement, the mucosa with muscularis mucosae and a small portion of upper submucosa should be available for histologic examination. These specimens should be evaluated in a systematic fashion, including assessment of (a) villous architecture, (b) surface and crypt epithelia, (c) lamina propria constituents, and (d) submucosal structures (53). A well-oriented specimen is essential for optimal evaluation. However, it must be remembered that villi vary in length and shape, particularly in the proximal duodenum, and that villous apices bend and twist in various planes to create unusual forms; these variations should not be misinterpreted as a villous abnormality (40,53). Generally, if four normal villi in a row are observed, the villous architecture of the entire specimen is probably normal (53,107). This does not mean that specimens with fewer than four well-aligned normal villi should be considered inadequate because even one normal intestinal villus in a proximal small bowel biopsy specimen rules out unrelated celiac sprue (31). Conversely, identification of four normal villi in a row does not necessarily exclude focal lesions, although it almost always does (107). The pathologist must be wary of certain common artifacts that may lead to erroneous interpretations. Careful attention to certain features (described below) within the various mucosal compartments will aid in their recognition.
Tangential Sectioning
Inappropriate orientation of the specimen, occurring at any point during processing, will lead to various tangential cuts or sections. The mucosa must be sectioned perpendicular to its long axis, or a distorted pattern disclosing apparently short and broad villi and an expanded lamina proprial compartment will be observed. However, several features aid in recognizing an oblique cut: (a) numerous elliptically shaped glands, (b) a multilayered arrangement of the crypts (Figure 24.35), or (c) a multilayered surface epithelium (53) (Figure 24.36). If any of these features are present, the villous architecture must be interpreted with caution.
Brunner's Gland Related Artifact
Brunner's glands have an inconsistent effect on villous architecture (31). Occasionally, normal length villi can be encountered overlying the Brunner's glands (Figure 24.37), but more commonly the villi appear distorted, short, broad, and stubby (31,40) (Figure 24.24). To minimize the potential effects of this artifact on interpretation, biopsy specimens of the small bowel for evaluation of malabsorptive states are routinely obtained as distally as possible in
P.623
the duodenum or from the proximal jejunum (i.e., near the ligament of Treitz) (76,134). Occasionally, however, more proximal small bowel biopsies are necessary for evaluation of duodenitis or ulcer disease.
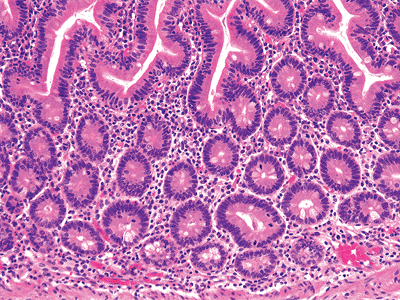 |
Figure 24.35 Multilayering of crypts indicates a tangential or oblique section of small intestine. The villous architecture overlying the crypts is normal, albeit unusual in appearance; this is also a product of malorientation of the biopsy specimen. |
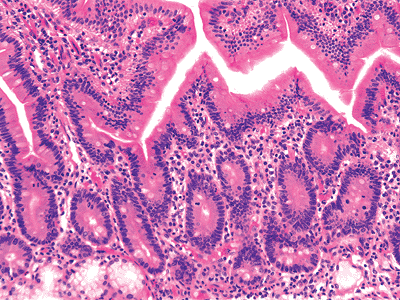 |
Figure 24.36 Another clue in identifying a tangential cut is multilayering, or stratification, of the surface epithelium (left portion of central villus). The normal surface layer is one-cell thick. This broad and short villous appearance is a consequence of malorientation and should be interpreted accordingly. |
Lymphoid Aggregate Related Artifact
Mucosal lymphoid aggregates, or nodules, are scattered along the small bowel and often distort the villous architecture. Villi are usually absent over lymphoid aggregates, and nearby villous forms may be distorted, short, and stubby (53) (Figure 24.31). Therefore, when a lymphoid aggregate is seen below an isolated flat portion of the surface epithelium, it should not be misinterpreted as a severe villous abnormality.
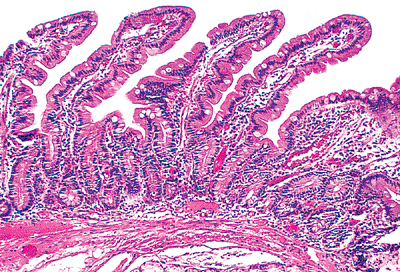 |
Figure 24.37 Occasionally, long slender villi, similar to those seen in the jejunum, are found overlying Brunner's glands. However, villi associated with Brunner's glands are more commonly shorter and broader (see Figure 24.24). |
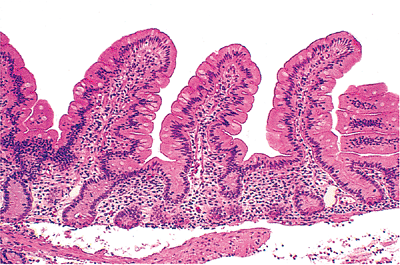 |
Figure 24.38 Absence of muscularis mucosae in a small bowel biopsy specimen, resulting in shorter- and broader-appearing villi that are widely spaced. |
Absence of Muscularis Mucosae
As mentioned earlier, the muscularis mucosae is an important structural component of the mucosa. In its absence (for instance, in a very superficial mucosal biopsy specimen), the tissues tend to spread laterally, resulting in villi becoming more widely spaced and appearing short and broad (53,135) (Figure 24.38).
Biopsy Trauma Related Artifacts
As a direct result of the traumatic pinch or suction biopsy procedures, certain alterations of normal mucosa can be seen. Separation of the villous surface epithelium from the underlying lamina propria or focally denuded epithelium are not unusual (53,135). The lack of acute erosive changes (e.g., neutrophilic infiltrate, cellular necrosis) or chronic evidence of ulceration (e.g., granulation tissue, regenerative epithelium) allow this alteration to be recognized as biopsy related. Additionally, focal hemorrhage and scattered polymorphonuclear leukocytes may be observed in the lamina propria as a consequence of the biopsy procedure. Crush or compression artifact can occur at the site of closure of the endoscopic forceps, resulting in a condensation of the lymphoplasmacytic component that can be misinterpreted as increased chronic inflammation (136). Additionally, the connective tissue may be altered in such a way that it appears more tightly packed, mimicking fibrosis or excessive collagen deposition (136).
P.624
Fixative-Related Artifacts
Certain fixatives other than formalin (4% formaldehyde solution) can cause interpretive problems. Although Hollande's fixative better preserves cytologic and nuclear detail, several artifacts may interfere with evaluation. The brightly eosinophilic granules of Paneth's cells and sometimes eosinophilic leukocytes seen readily in formalin-fixed tissue are not as well preserved by Hollande's fixative. Additionally, suboptimal clearing of Hollande's fixative from the specimens before paraffin embedding can result in residual minute, round, basophilic structures that resemble yeast forms or parasites (e.g., cryptosporidium, Giardia lamblia) (136)
Acknowledgment
We thank James T. McMahon, Ph.D., for the scanning electron micrographs.
References
1. Williams PL, Warwick R, Dyson M, Bannister LH, eds. Gray's Anatomy. 37th ed. New York: Churchill Livingstone; 1989.
2. Thorek P. Anatomy and Surgery. 3rd ed. New York: Springer-Verlag; 1985.
3. Costacurta L. Anatomical and functional aspects of the human suspensory muscle of the duodenum. Acta Anat (Basel) 1972;82:34 46.
4. Trier JS, Winter HS. Anatomy, embryology and developmental abnormalities of the small intestine and colon. In: Sleisenger MH, Fordtran JS, eds. Gastrointestinal Disease: Pathophysiology, Diagnosis, and Management. 4th ed. Philadelphia: WB Saunders; 1989:991 1021.
5. Parks DA, Jacobson ED. Physiology of the splanchnic circulation. Arch Intern Med 1985;145:1278 1281.
6. Alpers DH. Digestion and absorption of carbohydrates and proteins. In: Johnson LR, ed. Physiology of the Gastrointestinal Tract. 2nd ed. New York: Raven Press; 1987:1469 1487.
7. Davenport HW. Physiology of the Digestive Tract. 5th ed. Chicago: Year Book Medical; 1982.
8. Quigley EM. Small intestinal motor activity its role in gut homeostasis and disease. Q J Med 1987;65:799 810.
9. Fiorenza V, Yee YS, Zfass AM. Small intestinal motility: normal and abnormal function. Am J Gastroenterol 1987;82:1111 1114.
10. Lewin KJ. The endocrine cells of the gastrointestinal tract: the normal endocrine cells and their hyperplasias. In: Sommers SC, Rosen PP, Fechner RE, eds. Pathology Annual. Part 1. Norwalk, CT: Appleton-Century-Crofts; 1986:1 27.
11. Solcia E, Capella C, Buffa R, Usellini L, Fiocca R, Sessa F. Endocrine cells of the digestive system. In: Johnson LR, ed. Physiology of the Gastrointestinal Tract. 2nd ed. New York: Raven Press; 1987:111 130.
12. Elson CO, Kagnoff MF, Fiocchi C, Befus AD, Targan S. Intestinal immunity and inflammation: recent progress. Gastroenterology 1986;91:746 768.
13. Rubin W. The epithelial membrane of the small intestine. Am J Clin Nutr 1971;24:45 64.
14. Wilson JP. Surface area of the small intestine in man. Gut 1967;8:618 621.
15. Holmes R, Hourihane DO, Booth CC. The mucosa of the small intestine. Postgrad Med J 1961;37:717 724.
16. Toner PG, Carr KE. The use of scanning electron microscopy in the study of the intestinal villi. J Pathol 1969;97:611 617.
17. Trier JS, Madara JL. Functional morphology of the mucosa of the small intestine. In: Johnson LR, ed. Physiology of the Gastrointestinal Tract. 2nd ed. New York: Raven Press; 1987:1209 1249.
18. Dobbins WO. The intestinal mucosal lymphatic in man. A light and electron microscopic study. Gastroenterology 1966;51:994 1003.
19. Golab B, Szkudlarek R. Lymphatic vessels of the duodenum deep network. Folia Morphol (Warsz) 1981;39:263 270.
20. Neutra MR, Padykula HK. The gastrointestinal tract. In: Weiss L, ed. Modern Concepts of Gastrointestinal Histology. New York: Elsevier; 1984:658 706.
21. Poley JR. Loss of the glycocalyx of enterocytes in small intestine: a feature detected by scanning electron microscopy in children with gastrointestinal intolerance to dietary protein. J Pediatr Gastroenterol Nutr 1988;7:386 394.
22. Trier JS. The surface coat of gastrointestinal epithelial cells. Gastroenterology 1969;56:618 622.
23. Dawson IMP. Atlas of gastrointestinal pathology as seen on biopsy. In: Gresham GA, ed. Current Histopathology. Vol 6. Philadelphia: JB Lippincott; 1983:63-67.
24. Filipe MI. Mucins in the human gastrointestinal epithelium: a review. Invest Cell Pathol 1979;2:195 216.
25. Dobbins WO III. Human intestinal intraepithelial lymphocytes. Gut 1986;27:972 985.
26. Ferguson A, Murray D. Quantitation of intraepithelial lymphocytes in human jejunum. Gut 1971;12:988 994.
27. Selby WS, Janossy G, Bofill M, Jewell DP. Lymphocyte subpopulations in the human small intestine: the findings in normal mucosa and in the mucosa of patients with adult coeliac disease. Clin Exp Immunol 1983;52:219 228.
28. Cerf-Bensussan N, Schneeberger EE, Bhan AK. Immunohistologic and immunoelectron microscopic characterization of the mucosal lymphocytes of human small intestine by the use of monoclonal antibodies. J Immunol 1983;130:2615 2622.
29. Greenwood JH, Austin LL, Dobbins WO III. In vitro characterization of human intestinal intraepithelial lymphocytes. Gastroenterology 1983;85:1023 1035.
30. Kagnoff MF. Immunology and disease of the gastrointestinal tract. In: Sleisenger MH, Fordtran JS, eds. Gastrointestinal Disease. 4th ed. Philadelphia: WB Saunders; 1989:114 144.
31. Petras RE. Nonneoplastic intestinal disease. In: Sternberg SS, ed. Diagnostic Surgical Pathology. Vol 2. Philadelphia: Lippincott Williams & Wilkin; 2004:1475 1542.
32. Ferguson A, Sutherland A, MacDonald TT, Allan F. Technique for microdissection and measurement in biopsies of human small intestine. J Clin Pathol 1977;30:1068 1073.
33. Lipkin M. Proliferation and differentiation of normal and diseased gastrointestinal cells. In: Johnson LR, ed. Physiology of the Gastrointestinal Tract. 2nd ed. New York: Raven Press; 1987:255 284.
34. Williamson RC. Intestinal adaptation (first of two parts). Structural, functional, and cytokinetic changes. N Engl J Med 1978;298:1393 1402.
35. Watson AJM. Necrosis and apoptosis in the gastrointestinal tract. Gut 1995;37:165 167.
36. Facer P, Bishop AE, Lloyd RV, Wilson BS, Hennessy RJ, Polak JM. Chromogranin: a newly recognized marker for endocrine cells of the human gastrointestinal tract. Gastroenterology 1985;89:1366 1373.
37. Albrecht S, Gardiner GW, Kovacs K, Ilse G, Kaiser U. Duodenal somatostatinoma with psammoma bodies. Arch Pathol Lab Med 1989;113:517 520.
38. Sjolund K, Sanden G, Hakanson R, Sundler F. Endocrine cells in human intestine: an immunocytochemical study. Gastroenterology 1983;85:1120 1130.
39. Walsh JH. Gastrointestinal peptide hormones. In: Sleisenger MH, Fordtran JS, eds. Gastrointestinal Disease. 4th ed. Philadelphia: WB Saunders; 1989:78 9107.
40. Goldman H, Antonioli DA. Mucosal biopsy of the esophagus, stomach, and proximal duodenum. Hum Pathol 1982;13:423 448.
41. Sandow MJ, Whitehead R. The Paneth cell. Gut 1979;20:420 431.
42. Geller SA, Thung SN. Morphologic unity of Paneth cells. Arch Pathol Lab Med 1983;107:476 479.
P.625
43. Shen B, Porter E, Reynoso E, Shen C, Ghosh D, Connor J, Drazba J, Rho H, Gramlich T, Li R, Ormsby A, Sy M, Ganz T, Bevins C. Human defensin -5 expression in intestinal metaplasia of the upper gastrointestinal tract. J Clin Path; 2005; 58:687 694.
44. Jenkins D, Goodall A, Scott BB. T-lymphocyte populations in normal and coeliac small intestinal mucosa defined by monoclonal antibodies. Gut 1986;27:1330 1337.
45. Lee FD, Toner PG. Biopsy Pathology of the Small Intestine. Philadelphia: JB Lippincott; 1980.
46. Isaacson P, Judd MA. Carcinoembryonic antigen (CEA) in the normal human small intestine: a light and electron microscopic study. Gut 1977;18:786 791.
47. Scott H, Solheim BG, Brandtzaeg P, Thorsby E. HLA-DR like antigens in the epithelium of the human small intestine. Scand J Immunol 1980;12:77 82.
48. Chiba M, Ohta H, Nagasaki A, Arakawa H, Masamune O. Lymphoid cell subsets in normal human small intestine. Gastroenterol Jpn 1986;21:336 343.
49. Brandtzaeg P, Halstensen TS, Kett K, et al. Immunobiology and immunopathology of human gut mucosa: humoral immunity and intraepithelial lymphocytes. Gastroenterology 1989;97:1562 1584.
50. Comer GM, Brandt LJ, Abissi CJ. Whipple's disease: a review. Am J Gastroenterol 1983;78:107 114.
51. Strom RL, Gruninger RP. AIDS with Mycobacterium avium-intracellulare lesions resembling those of Whipple's disease. N Engl J Med 1983;309:1323 1325.
52. Roth RI, Owen RL, Keren DF, Volberding PA. Intestinal infection with Mycobacterium avium in acquired immune deficiency syndrome (AIDS). Histological and clinical comparison with Whipple's disease. Dig Dis Sci 1985;30:497 504.
53. Perera DR, Weinstein WM, Rubin CE. Symposium on pathology of the gastrointestinal tract. Part II. Small intestinal biopsy. Hum Pathol 1975;6:157 217.
54. Klein NC, Hargrove RL, Sleisenger MH, Jeffries GH. Eosinophilic gastroenteritis. Medicine (Baltimore) 1970;49:299 319.
55. Johnstone JM, Morson BC. Eosinophilic gastroenteritis. Histopathology 1978;2:335 348.
56. Goldman H, Proujansky R. Allergic proctitis and gastroenteritis in children. Clinical and mucosal biopsy features in 53 cases. Am J Surg Pathol 1986;10:75 86.
57. DeSchryver-Kecskemeti K, Clouse RE. A previously unrecognized subgroup of eosinophilic gastroenteritis. Association with connective tissue diseases. Am J Surg Pathol 1984;8:171 180.
58. McNabb PC, Fleming CR, Higgins JA, Davis GL. Transmural eosinophilic gastroenteritis with ascites. Mayo Clin Proc 1979;54:119 122.
59. Heatley RV. The gastrointestinal mast cell. Scand J Gastroenterol 1983;18:449 453.
60. Befus D, Goodacre R, Dyck N, Bienenstock J. Mast cell heterogeneity in man. I. Histologic studies of the intestine. Int Arch Allergy Appl Immunol 1985;76:232 236.
61. Scott BB, Hardy GJ, Losowsky MS. Involvement of the small intestine in systemic mast cell disease. Gut 1975;16:918 924.
62. Lundqvist M, Wilander E. Subepithelial neuroendocrine cells and carcinoid tumors of the human small intestine and appendix. A comparative immunohistochemical study with regard to serotonin, neuron-specific enolase and S-100 protein reactivity. J Pathol 1986;148:141 147.
63. Lord MG, Valies P, Broughton AC. A morphologic study of the submucosa of the large intestine. Surg Gynecol Obstet 1977;145:55 60.
64. Lee AK, DeLellis RA, Silverman ML, Wolfe HJ. Lymphatic and blood vessel invasion in breast carcinoma: a useful prognostic indicator? Hum Pathol 1986;17:984 987.
65. Schlingemann RO, Dingjan GM, Emeis JJ, Blok J, Warnaar SO, Ruiter DJ. Monoclonal antibody PAL-E specific for endothelium. Lab Invest 1985;52:71 76.
66. Vardy PA, Lebenthal E, Shwachman H. Intestinal lymphagiectasia: a reappraisal. Pediatrics 1975;55:842 851.
67. Gershon MD, Erde SM. The nervous system of the gut. Gastroenterology 1981;80:1571 1594.
68. Goyal RK, Crist JR. Neurology of the gut. In: Sleisenger MH, Fordtran JS, eds. Gastrointestinal Disease. 4th ed. Philadelphia: WB Saunders; 1989:21 52.
69. Krishnamurthy S, Schuffler MD. Pathology of neuromuscular disorders of the small intestine and colon. Gastroenterology 1987;93:610 639.
70. Ferri GL, Probert L, Cocchia D, Michetti F, Marangos PJ, Polak JM. Evidence for the presence of S-100 protein in the glial component of the human enteric nervous system. Nature 1982;297:409 410.
71. Romert P, Mikkelsen HB. cC-kit immunoreactive interstitial cells of Cajal in the human small and large intestine. Histochem Cell Biol 1998; 109:195 202.
72. Lawson HH. The duodenal mucosa in health and disease. A clinical and experimental study. Surg Annu 1989;21:157 180.
73. Lawson HH. Definition of the gastroduodenal junction in healthy subjects. J Clin Pathol 1988;41:393 396.
74. Korn ER, Foroozan P. Endoscopic biopsies of normal duodenal mucosa. Gastrointest Endosc 1974;21:51 54.
75. Kreuning J, Bosman FT, Kuiper G, Wal AM, Lindeman J. Gastric and duodenal mucosa in healthy individuals. An endoscopic and histopathological study of 50 volunteers. J Clin Pathol 1978;31:69 77.
76. Dandalides SM, Carey WD, Petras RE, Achkar E. Endoscopic small bowel mucosal biopsy: a controlled trial evaluating forceps size and biopsy location in the diagnosis of normal and abnormal mucosal architecture. Gastrointest Endosc 1989;35:197 200.
77. Robertson HE. The pathology of Brunner's glands. Arch Pathol 1941;31:112 130.
78. Lang IM, Tansy MF. Brunner's glands. In: Young JA, ed. Gastrointestinal Physiology. IV. International Review of Physiology. Vol 28. Baltimore: University Park Press; 1983:85 102.
79. Treasure T. The ducts of Brunner's glands. J Anat 1978;127:299 304.
80. Leeson TS, Leeson RC. The fine structure of Brunner's glands. J Anat 1968;103:263 276.
81. Thompson IW, Day DW, Wright NA. Subnuclear vacuolated mucous cells: a novel abnormality of simple mucin-secreting cells of non-specialized gastric mucosa and Brunner's glands. Histopathology 1987;11:1067 1081.
82. Kamiya R. Basal-granulated cells in human Brunner's glands. Arch Histol Jpn 1983;46:87 101.
83. Bosshard A, Chery-Croze S, Cuber JC, Dechelette MA, Berger F, Chayvialle JA. Immunocytochemical study of peptidergic structures in Brunner's glands. Gastroenterology 1989;97:1382 1388.
84. Silverman L, Waugh JM, Huizenga KA, Harrison EG Jr. Large adenomatous polyp of Brunner's glands. Am J Clin Pathol 1961;36:438 443.
85. Franzin G, Musola R, Ghidini O, Manfrini C, Fratton A. Nodular hyperplasia of Brunner's glands. Gastrointest Endosc 1985;31:374 378.
86. West B. Pseudomelanosis duodeni. J Clin Gastroenterol 1988; 10:127 129.
87. Rosenberg JC, DiDio LJ. Anatomic and clinical aspects of the junction of the ileum with the large intestine. Dis Colon Rectum 1970;13:220 224.
88. Kumar D, Phillips SF. The contribution of external ligamentous attachments to function of the ileocecal junction. Dis Colon Rectum 1987;30:410 416.
89. Axelsson C, Andersen JA. Lipohyperplasia of the ileocaecal region. Acta Chir Scand 1974;140:649 654.
90. Cornes JS. Number, size, and distribution of Peyer's patches in the human small intestine. Part I. The development of Peyer's patches. Part II. The effect of age on Peyer's patches. Gut 1965;6:225 233.
91. MacDonald TT, Spencer J, Viney JL, Williams CB, Walker-Smith JA. Selective biopsy of human Peyer's patches during ileal endoscopy. Gastroenterology 1987;93:1356 1362.
92. Spencer J, Finn T, Isaacson PG. Human Peyer's patches: an immunohistochemical study. Gut 1986;27:405 410.
93. Pang LC. Intussusception revisited: clinicopathologic analysis of 261 cases with emphasis on pathogenesis. South Med J 1989;82:215 228.
P.626
94. Schenken JR, Kruger RL, Schultz L. Papillary lymphoid hyperplasia of the terminal ileum: an unusual cause of intussusception and gastrointestinal bleeding in childhood. J Pediatr Surg 1975;10:259 265.
95. Fieber SS, Schaefer HJ. Lymphoid hyperplasia of the terminal ileum a clinical entity? Gastroenterology 1966;50:83 98.
96. Bjerke K, Brandtzaeg P, Fausa O. T cell distribution is different in follicle-associated epithelium of human Peyer's patches and villous epithelium. Clin Exp Immunol 1988;74:270 275.
97. Wolf JL, Bye WA. The membranous epithelial (M) cell and the mucosal immune system. Annu Rev Med 1984;35:95 112.
98. Owen RL, Jones AL. Epithelial cell specialization within human Peyer's patches: an ultrastructural study of intestinal lymphoid follicles. Gastroenterology 1974;66:189 203.
99. Shepherd NA, Crocker PR, Smith AP, Levison DA. Exogenous pigment in Peyer's patches. Hum Pathol 1987;18:50 54.
100. Urbanski SJ, Arsenault AL, Green FH, Haber G. Pigment resembling atmospheric dust in Peyer's patches. Mod Pathol 1989;2:222 226.
101. Mackey WC, Dineen P. A fifty year experience with Meckel's diverticulum. Surg Gynecol Obstet 1983;156:56 64.
102. Artigas V, Calabuig R, Badia F, Rius X, Allende L, Jover J. Meckel's diverticulum: value of ectopic tissue. Am J Surg 1986;151:631 634.
103. Bennett MK, Sachdev GK, Jewell DP, Anand BS. Jejunal mucosal morphology in healthy north Indian subjects. J Clin Pathol 1985;38:368 371.
104. Cook GC, Kajubi SK, Lee FD. Jejunal morphology of the African in Uganda. J Pathol 1969;98:157 169.
105. Lindenbaum J, Gerson CD, Kent TH. Recovery of small-intestinal structure and function after residence in the tropics. I. Studies in Peace Corps volunteers. Ann Intern Med 1971;74:218 222.
106. Gerson CD, Kent TH, Saha JR, Siddiqi N, Lindenbaum J. Recovery of small-intestinal structure and function after residence in the tropics. II. Studies in Indians and Pakistanis living in New York City. Ann Intern Med 1971;75:41 48.
107. Dobbins WO III. Small bowel biopsy in malabsorptive states. In: Norris HT, ed. Pathology of the Colon, Small Intestine, and Anus. New York: Churchill Livingstone; 1983:121 167.
108. Webster SG, Leeming JT. The appearance of the small bowel mucosa in old age. Age Ageing 1975;4:168 174.
109. Chacko CJ, Paulson KA, Mathan VI, Baker SJ. The villus architecture of the small intestine in the tropics: a necropsy study. J Pathol 1969;98:146 151.
110. Wolff M. Heterotopic gastric epithelium in the rectum: a report of three new cases with a review of 87 cases of gastric heterotopia in the alimentary canal. Am J Clin Pathol 1971;55:604 616.
111. Lessells AM, Martin DF. Heterotopic gastric mucosa in the duodenum. J Clin Pathol 1982;35:591 595.
112. Tsubone M, Kozuka S, Taki T, Hoshino M, Yasui A, Hachisuka K. Heterotopic gastric mucosa in the small intestine. Acta Pathol Jpn 1984;34:1425 1431.
113. Lai EC, Tompkins RK. Heterotopic pancreas. Review of a 26 year experience. Am J Surg 1986;151:697 700.
114. Dolan RV, ReMine WH, Dockerty MB. The fate of heterotopic pancreatic tissue. A study of 212 cases. Arch Surg 1974;109:762 765.
115. Armstrong CP, King PM, Dixon JM, Macleod IB. The clinical significance of heterotopic pancreas in the gastrointestinal tract. Br J Surg 1981;68:384 387.
116. Tanemura H, Uno S, Suzuki M, et al. Heterotopic gastric mucosa accompanied by aberrant pancreas in the duodenum. Am J Gastroenterol 1987;82:685 688.
117. Tomasi TB Jr. Mechanisms of immune regulation at mucosal surfaces. Rev Infect Dis 1983;5(suppl 4):S784 S792.
118. Ranchod M, Lewin KJ, Dorfman RF. Lymphoid hyperplasia of the gastrointestinal tract: a study of 26 cases and review of the literature. Am J Surg Pathol 1978;2:383 400.
119. Rambaud JC, DeSaint-Louvent P, Marti R, et al. Diffuse follicular lymphoid hyperplasia of the small intestine without primary immunoglobulin deficiency. Am J Med 1982;73:125 132.
120. Matuchansky C, Touchard G, Lemaire M, et al. Malignant lymphoma of the small bowel associated with diffuse nodular lymphoid hyperplasia. N Engl J Med 1985;313:166 171.
121. Hermans PE, Diaz-Buxo JA, Stobo JD. Idiopathic late-onset immunoglobulin deficiency: clinical observations in 50 patients. Am J Med 1976;61:221 237.
122. Lewin KJ, Kahn LB, Novis BH. Primary intestinal lymphoma of Western and Mediterranean type, alpha chain disease and massive plasma cell infiltration: a comparative study of 37 cases. Cancer 1976;38:2511 2528.
123. Lewin KJ, Ranchod M, Dorfman RF. Lymphomas of the gastrointestinal tract: a study of 117 cases presenting with gastrointestinal disease. Cancer 1978;42:693 707.
124. Grody WW, Magidson JG, Weiss LM, Hu E, Warnke HA, Lewin KJ. Gastrointestinal lymphomas: immunohistochemical studies on the cell of origin. Am J Surg Pathol 1985;9:328 337.
125. Tubbs RR, Sheibani K. Immunohistology of lymphoproliferative disorders. Semin Diagn Pathol 1984;1:272 284.
126. Little JV, Foucar K, Horvath A, Crago S. Flow cytometric analysis of lymphoma and lymphoma-like disorders. Semin Diagn Pathol 1989;6:37 54.
127. Grody WW, Gatti RA, Naiem F. Diagnostic molecular pathology. Mod Pathol 1989; 2:553 568.
128. Goldman H, Antonioli DA. Mucosal biopsy of the rectum, colon, and distal ileum. Hum Pathol 1982;13:981 1012.
129. Bechi P, Romagnoli P, Cortesini C. Ileal mucosal morphology after total colectomy in man. Histopathology 1981;5:667 678.
130. Philipson B, Brandberg A, Jagenburg R, Kock NG, Lager I, Ahren C. Mucosal morphology, bacteriology, and absorption in intra-abdominal ileostomy reservoir. Scand J Gastroenterol 1975;10:145 153.
131. Shepherd NA, Jass JR, Duval I, Moskowitz RI, Nicholls RJ, Morson BC. Restorative proctocolectomy with ileal reservoir: pathological and histochemical study of mucosal biopsy specimens. J Clin Pathol 1987;40:601 607.
132. Trier JS. Diagnostic value of peroral biopsy of the proximal small intestine. N Engl J Med 1971;285:1470 1473.
133. Brandborg LL, Rubin GE, Quinton WE. A multipurpose instrument for suction biopsy of the esophagus, stomach, small bowel, and colon. Gastroenterology 1959;37:1 16.
134. Achkar E, Carey WD, Petras R, Sivak MV, Revta R. Comparison of suction capsule and endoscopic biopsy of small bowel mucosa. Gastrointest Endosc 1986;32:278 281.
135. Whitehead R. Mucosal biopsy of the gastrointestinal tract. In: Bennington JL, ed. Major Problems in Pathology. Vol 3. 3rd ed. Philadelphia: WB Saunders; 1985.
136. Haggitt RC. Handling of gastrointestinal biopsies in the surgical pathology laboratory. Lab Med 1982;13:272 278.