6 - Myofibroblast
Editors: Mills, Stacey E.
Title: Histology for Pathologists, 3rd Edition
Copyright 2007 Lippincott Williams & Wilkins
> Table of Contents > III - Musculoskeletal System > 9 - Blood Vessels
function show_scrollbar() {}
9
Blood Vessels
Patrick J. Gallagher
Allard C. van der Wal
Gross and Light Microscopic Features
The normal structure of vessels, particularly the aorta, elastic and muscular arteries, and the larger veins, change progressively throughout life (Table 9.1) (1,2). These aging changes lead to increased arterial stiffness, detected clinically by alterations in pulse wave velocity (3). It is now clear that aging arteries are especially affected by common disorders such as atherosclerosis, hypertension, and diabetes (Table 9.2). Surgical pathologists must be fully aware not only of the nature and extent of these alterations, but also of their variation from site to site.
Aorta
The length and breadth of the aorta increase progressively throughout life. Although there are some variations in the rate of these changes, both between men and women and from decade to decade, the process continues well into a person's seventies and eighties. This enlargement produces the characteristic unfolding of the aorta so often seen in chest radiographs; and, if the aortic valve annulus is also involved, aortic incompetence can result. Some atherosclerosis is almost inevitable in the abdominal aorta in the middle aged and elderly, but aging changes are independent of this.
The principal components of all arteries are elastic and collagen fibers, smooth muscle cells, and a mucopolysaccharide-rich ground substance (4). In the media of the aorta and the carotid, the innominate and proximal axillary arteries elastic fibers predominate. Parallel lamellar units of elastin enclose smooth muscle cells, ground substance, and collagen (Figure 9.1). There are about 40 lamellar units at birth and at least 50 in adult life, each measuring about 11 m in thickness. Interconnecting bands of collagen and elastin fibers provide strength, whereas the lamellar arrangement distributes stress evenly across the wall, smoothing the cyclical pressure waves of cardiac contraction (5). Elastic fragmentation with associated foci of fibrosis are the most prominent aging changes and account for the weakening that leads to aortic dilatation (Figure 9.2). The changes in the structure of the extracellular matrix are
P.218
thought to be the result of upregulation of genes that mediate matrix metalloproteinase production (6). Apoptosis can be demonstrated in a number of cell types within atheromatous plaques (7) but is unlkely to be a key factor in the aging of the arterial wall (8). Calcification is a common complication; and, although it is most frequent in atheromatous segments, it may occur in areas where the intima is virtually normal. The underlying causes of aortic and coronary arterial calcification remain poorly understood (9,10). Small amounts of amyloid can be detected in aortic atheromatous lesions of middle-aged and elderly subjects and may be derived from serum amyloid A or other apolipoproteins (11,12).
Table 9.1 Aging Changes in Blood Vessels | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
Cystic medial degeneration (CMD), originally called medionecrosis aortae by Erdheim, is a difficult concept, and many pathologists are unsure about the exact meaning of the term. Histologically, the condition is characterized by degeneration and fragmentation of the elastic layers of the media and formation of mucoid pools (Figure 9.1B C). Some areas have few, if any, stainable nuclei, and this is the result of smooth muscle cell death. More recently, areas of smooth muscle cell apoptosis and disorganized proliferation, fibrosis, and angiogenesis have been described, suggesting that CMD is a process of degenerative injury and repair. In 1977, Schlatmann and Becker (13) showed that the histological alterations of CMD showed a striking correlation with age and may therfore represent the normal aging process of elastic arteries. The same features are seen in hypertensive patients, who have an altered hemodynamic profile, and in genetic disorders of connective tissue, such as Marfan's or some types of the Ehlers-Danlos syndrome. They have also been reported in patients with a history of cocaine abuse (14). In connective tissue disorders, CMD is more pronounced and leads to complications such as intramural hematoma formation or aortic dissection at an earlier age.
Although the exact cause of CMD is unknown, it appears to be related to an imbalance between the mechanical forces imposed on the aortic wall during systole and the
P.219
P.220
capacity of the aortic wall to resist these forces. The resulting shear forces may cause alterations in the secretion pattern of smooth muscle cells or their death by apoptosis. More recently, p53 accumulation, bax upregulation, and both vascular smooth muscle cell apoptosis and regeneration have been demonstrated in areas of cystic medial necrosis (15). In Marfan's syndrome, the histologic changes suggest exaggerated aging, but there are no features that allow a specific diagnosis to be made (Figure 9.2B). The underlying genetic abnormality involves a glycoprotein, fibrillin, that is closely associated with elastin fibers. The exact functions of fibrillin and other associated glycoproteins are uncertain, but they may act as a scaffold on which elastin fibers are laid down. There is a wide spectrum of clinical abnormalities in Marfan's syndrome, and certain clinical features, such as arachnodactyly or aortic dissection, are especially common in some families (16).
Table 9.2 Histologic Changes in Arteries and Arterioles | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
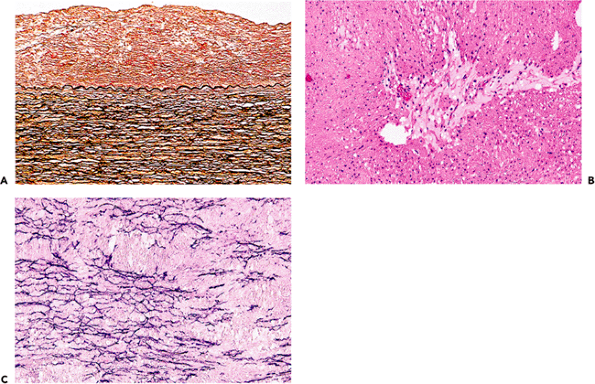 |
Figure 9.1 A. Inner half of the aortic wall of a 62-year-old man. There is a moderate degree of fibrous intimal thickening, which has no immediate clinical relevance but may predispose to atherosclerosis. There was only slight fragmentation of the elastic lamellae; the overall appearance is well within normal limits for a patient of this age (elastic van Gieson). B. The typical appearance of cystic medial degeneration in an H&E stained section. Note the prominent pool of mucoid material. C. This shows a corresponding section to B but is stained for elastic tissue. There is extensive loss of the normal elastic framework. |
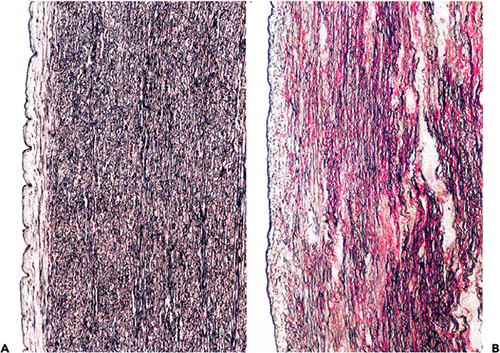 |
Figure 9.2 A. The normal appearance of the aortic media of a 48-year-old man. There are many parallel lamellae of elastic tissue. There is no significant intimal thickening. B. The aortic wall of a 31-year-old man with Marfan's syndrome. The medial elastic tissue is extensively fragmented, and there is fibrosis and loose mucopolysaccharide-rich areas. Such extensive changes would be unusual even in an elderly patient. (Elastic van Gieson.) |
Elastic fragmentation and associated medial necrosis are the most common histologic findings in both ascending and thoracic aortic aneurysms. Despite thinning of the wall due to vascular dilatation, the cellular and matrix components in thoracic aneurysms are in fact increased as a result of vascular smooth muscle hyperplasia. In contrast in abdominal aortic aneurysms there is a reduction in smooth muscle density in abdominal aortic aneurysms (17,18). Traditionally, abdominal aortic aneurysms have been considered atheromatous in origin, but this is an oversimplification. Genetic studies have provided compelling evidence for an inherited basis of this disease. Aneurysms have been detected in up to 20% of first degree relatives, especially when the affected subject is female (19). Whether the atherosclerosis is the primary cause or a secondary complication, the inflammation and medial scarring that accompany all but the earliest stages of atheroma further damage a wall already weakened by normal aging or by specific genetically determined alterations in the matrix of the aortic wall. Patchy chronic inflammatory aggregates, including lymphocytes and plasma cells, are often present in the adventitia of atheromatous segments of the aorta and coronary arteries (Figure 9.3). In biopsies of the ascending aorta during repair of dissecting aneurysms or aortic reconstructions for root dilatation, these chronic adventitial infiltrates must not be mistaken as evidence of aortitis. Small collections of lymphocytes, macrophages, and giant cells are occasionally seen in these biopsies. Although they should be reported, our experience is that they have no clinical significance. In some abdominal aneurysms, the inflammatory infiltrates are especially dense, and surgical repair may be difficult. The inflammation may be a reaction to ceroid pigment, and there can be associated retroperitoneal fibrosis (20).
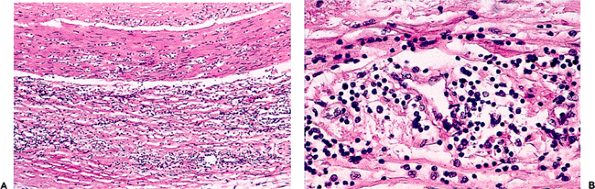 |
Figure 9.3 A. and B. Adventitial chronic inflammatory infiltrates in the wall of an atheromatous coronary artery. A few inflammatory cells have infiltrated into the media. The magnified view on the right confirms that most of the inflammatory cells are lymphocytes or plasma cells. |
Cardiac surgeons have several techniques for repairing aortic coarctations and may submit samples of aorta, the narrowed aortic segment, the subclavian artery, or the ductus arteriosus (arterial duct) for histologic identification. The aorta around the coarctation may show reactive intimal thickening, even in neonates, but the underlying elastic structure is usually well preserved. The coarctation itself can have a variety of appearances. In long-standing cases, there may be dense intimal and medial fibrosis. In neonates, the intima may have a distinctly irregular pattern of fibroelastic intimal thickening, resembling some forms of arterial dysplasia (Figure 9.4). The structure of the arterial duct changes progressively during intrauterine growth and in the postnatal period (21) and can be influenced by prostaglandin treatment. Unlike the aorta and the proximal subclavian artery, which are elastic vessels, the arterial duct has a muscular media and a defined internal elastic lamella.
Arteries
It is only in children and young adults that muscular arteries conform to the classical descriptions of textbooks. The
P.221
intima of arteries is defined as the region from and including the endothelium to the luminal margin of the media (22). At birth, the intima is a virtual space with the endothelium closely opposed to the internal elastic lamella. This layer thickens slowly with age, either (a) eccentrically at branching points or bifurcations or (b) diffusely. Both types occur preferentially at sites of altered blood flow or mechanical stress, suggesting that they are adaptive changes (a response to injury). Vascular smooth muscle cells derived from the underlying media and extracellular matrix proteins accumulate in the thickened intima and may serve as a soil for the development of atherosclerotic plaques. For example, in the aorta and coronary arteries, the so-called atherosclerosis prone areas are those that show early diffuse or eccentric thickening.
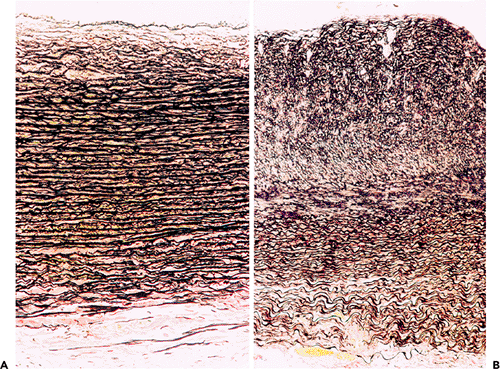 |
Figure 9.4 Coarctation of the aorta. A. The aortic wall distal to a coarctation in a 3-month-old child. There is slight intimal edema only. B. The coarctation itself; note the irregular arrangement of the intimal fibroelastic tissue. (Elastic van Gieson.) |
Progressive intimal fibrosis affects nearly all arteries (Figure 9.5), but in surgical pathology material it is especially noticeable in the spleen, myometrium, and thyroid (Figure 9.6). As in the aorta, fragmentation of the elastic tissue, usually the internal elastic lamella, is common and is of no specific significance (Figure 9.7). In some aging arteries, the internal lamella appears to repeatedly reduplicate, producing a pattern of concentric intimal thickening (Figure 9.5). Small foci of calcification can be identified in otherwise normal vessels, usually just to the medial aspect of the internal elastic lamella. These aging changes, often loosely termed arteriosclerosis, have been studied most extensively in the coronary arteries, where women generally show substantially less elastic fragmentation and intimal fibrosis than do men of the same age (45). About 75% of the mass of the media is smooth muscle cells. These run in a spiral or circumferential pattern around the wall. As in the intima the small amounts of associated collagen and elastin increase throughout life. Arteries dilate and become more tortuous with increasing age, and this has a fortuitous antiocclusive effect. The caliber of the coronary arteries in middle-aged and elderly women is less than that of men. This may make coronary artery surgery more difficult and contribute to the poorer results recorded in women (23). If arterial dilatation is pronounced and irregular, as in so-called coronary artery ectasia, spontaneous thrombosis may result.
Nutrients reach the media of elastic or muscular arteries by direct diffusion through the intima or via small branches, the vasa vasorum, which reenter the media from the adventitial aspect. Vasa are best seen in biopsy samples taken from the ascending aorta during root repairs and sometimes have remarkably thick muscular walls (Figure 9.8). In
P.222
P.223
atheromatous arteries, there is often marked neovascular proliferation. Hemorrhage from these vessels contributes to the growth of lesions and their lipid content (24).
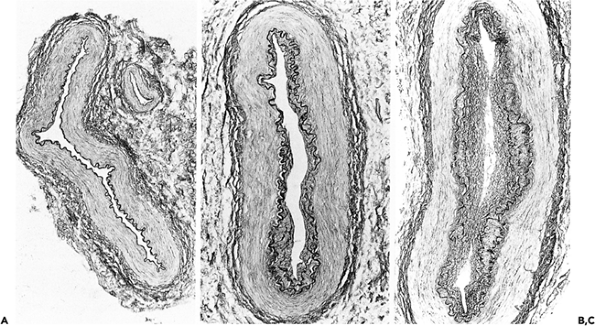 |
Figure 9.5 Aging changes in muscular arteries. A. (left) Normal appearing artery from a 17-year-old girl. B. and C. Note the progressive intimal fibrosis in arteries from elderly males. In C there is some reduplication of the internal elastic membrane. (Elastic van Gieson.) |
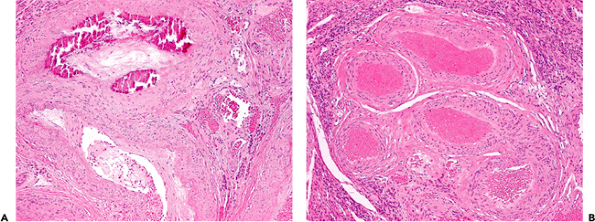 |
Figure 9.6 A. and B. Aging changes in arteries. These thick-walled vessels were close to the serosa of the myometrium in a 52-year-old woman. Note the prominent calcification in A and the increased tortuosity in B. These changes have no importance. They can be seen in other sites, especially in thyroidectomy specimens. |
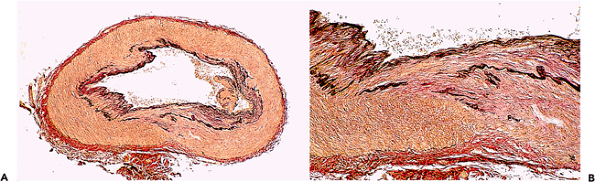 |
Figure 9.7 A. and B. Temporal artery from a 72-year-old woman who died suddenly from coronary heart disease. There was no past history of headache or temporal arteritis. Note the fragmentation of the elastic lamella with a little associated fibrosis (red coloration in B). Changes such as these are commonplace in the elderly and must not be interpreted as evidence of previous arteritis (elastic van Gieson). |
The different stages, or phases, of atherosclerotic plaque development have been categorized in the American Heart Association (AHA) classification into three types of early lesions (plaque types I III) and three types of mature late lesions (plaque types IV VI). Early lesions consist of adaptive intimal thickening (type I), fatty streaks (type II) with accumulation of lipid-rich foam cells and T lymphocytes), and pathological intimal thickening (type III, early atheroma). Late stages include fibroatheroma (type IV), fibrotic or calcified plaques (type V), and complicated plaques (type VI) (25,26). Recently, there has been much interest in the concept of the so-called vulnerable plaque: plaques at high risk for development of superimposed thrombosis or plaque hemorrhage. Several postmortem studies of coronary arteries from patients with myocardial infarction or sudden cardiac death have shown that vulnerable lesions have specific features such as a large lipid core, a thin fibrous cap, and marked inflammatory activity in the plaque tissue (27).
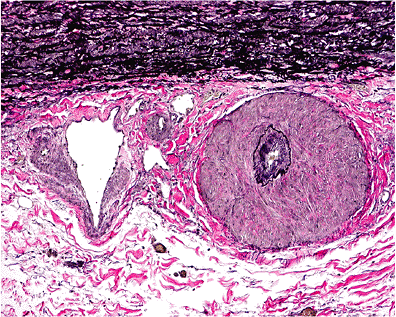 |
Figure 9.8 The aortic adventitia. The thick-walled vessel is a vasa vasorum. The thin-walled vessel (left) is a small vein. |
Virmani et al. (28) have modified the AHA classification in order to highlight the variation in plaque morphology in relation to the onset and evolution of atherothrombotic complications. In their classification, the late stages of plaque development are divided into fibrous cap atheroma, thin cap fibroatheroma, healed plaque rupture or erosion, and calcified plaques. This classification emphasizes that complicated (thrombosed) plaques can be the result of rupture of a fibrous cap or erosion of the endothelial lining (Table 9.3).
Inflammatory Infiltrates in Arteries
Apart from a few scattered macrophages or mast cells, the adventitia of arteries is devoid of inflammatory cells. However, for many years it has been known that, in the chronic advanced stages of atherosclerosis, nodular or patchy inflammatory infiltrates can form at the sites of atheromatous lesions, a process that increases with the severity of atherosclerosis (29). These aggregates can resemble the lymphoid follicle-like lesions that form in diseases of disordered immunity, such as rheumatoid disease. Immunohistochemistry has shown that these highly organized structures containing germinal centers surrounded by both T and B cells. Small vessels are lined by plump endothelium, which stains with antibodies that identify high endothelial venules (see Immunohistochemistry section,
P.224
below). Arteries in chronically inflamed tissues and within tumors often show pronounced fibrous intimal thickening, sometimes termed endarteritis obliterans (Table 9.2). In the early stages of this process, the fibrous tissue has a loose histologic appearance, and the ground substance may be basophilic. Although inflammatory or tumor cells often closely surround the adventitia, they do not usually penetrate far into the muscular wall.
Table 9.3 Modified American Heart Association Classification of Atheromatous Lesions | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ||||||||||||||||
These changes must be carefully distinguished from those of systemic vasculitis. In general terms, vasculitis tends to affect vessels of a specific size, cause necrosis of vessel walls (Figure 9.9) with associated hemorrhage, and lead to tissue infarction. In healed vasculitis, there is irregular fibrosis of the muscular wall (Figure 9.10).
Arterioles
There are no specific histologic features that accurately distinguish small arteries from larger arterioles; but, for convenience, arterioles are said to have a diameter of less than 100 m. However, in biopsy material, there is so much variation in the contours of these small vessels that accurate distinction is often impossible and probably unnecessary. Larger arterioles have an obvious media and an adventitial layer of connective tissue. In the smallest (terminal) arterioles, an internal elastic lamella may not be identified. The smooth muscle cells are arranged circumferentially, each cell winding around the wall several times. This is the structural basis of the precapillary sphincter. Small arterioles have a very thin adventitia but are richly supplied by sympathetic nerve fibers.
Hyalinization is a common lesion of arterioles and small arteries and increases with age and in conditions such as hypertension and diabetes. The glassy uniform appearance is the result of accumulation of a variety of plasma proteins and small amounts of lipids. As in arteries, reduplication of elastic tissue and intimal fibrosis are common changes in the aged. In severe longstanding benign hypertension and in the malignant phase, the arteriolar lumen can be substantially narrowed by concentric layers of fibrous tissue and smooth muscle cells, changes that are outside the normal range of aging (Table 9.1). Fibrinoid necrosis of the arteriolar media is the hallmark of malignant hypertension and some forms of acute vasculitis (Figure 9.9). It must always be regarded as pathologic. In the earliest changes of diabetic microangiopathy, arterioles and capillaries often show prominent basement membrane thickening (30). This thickening can be readily identified in renal and peripheral nerve biopsies. Although there is physiologic evidence of small vessel disease in the heart and the peripheral vasculature, characteristic histologic changes of diabetic microangiopathy are seldom seen in these sites. In diabetes the amounts of type IV collagen and laminin are increased, but the proteoglycan component of the basement membrane is reduced. Albumin and immunoglobulins accumulate in these abnormal basement membranes, binding to glycosylated protein residues and contributing to the overall eosinophilic appearance.
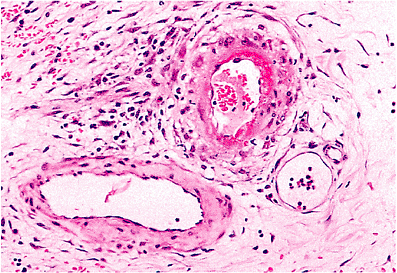 |
Figure 9.9 Florid fibrinoid necrosis in a small intestinal vessel of a girl with systemic lupus erythematosus. Fibrinoid necrosis is not a feature of normal aging or uncomplicated hypertension. It always should be regarded as pathologic. In this case, the involved vessel is probably an arteriole. Note the small vein (lower left) and capillary (lower right). |
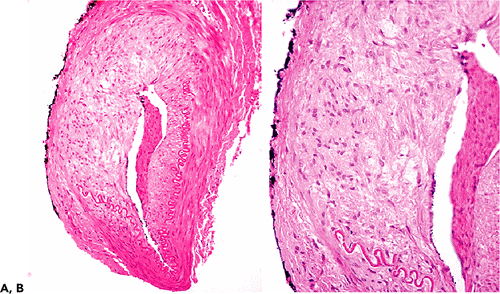 |
Figure 9.10 Healed temporal arteritis. This patient had been receiving steroid therapy for two weeks when this biopsy was performed. A. The low power view shows irregular thickening of the wall and a loss of about 50% of the internal elastic lamella. B. This higher power view shows fresh fibrous scarring of the media. Changes such as these are not part of the normal aging process (see Figure 9.6). |
P.225
Capillaries
In contrast to arterioles, capillaries have neither a muscular media nor an elastic lamella. A single but complete layer of endothelial cells lies on a basement membrane whose thickness varies from site to site. Basement membrane thickness increases with age, almost doubling in muscle capillaries from 10 to 70 years of age. There is no fibrous tissue support peripheral to this, but pericytes are present in and among the basement membrane. It is difficult to identify pericytes in routine sections, but they are easily seen by electron microscopy and also stain with antismooth muscle actin antibodies. They provide structural support; and, because they contain several forms of myosin, they may be able to regulate blood flow. It is likely that they are involved in the synthesis of vascular basement membrane and are capable of phagocytosis (31). It is generally accepted that the turnover of pericytes is increased in the capillaries of diabetics, and this may contribute to the development of small vessel disease (30).
The endothelium of capillaries may have circular fenestrations that act as pores through the full thickness of the endothelial cell. Fenestrations are especially prominent in renal glomerular endothelial cells and are found in the intestinal mucosa, skin, and endocrine glands. In contrast, fenestrations are poorly developed or absent in brain, muscle, lung, and connective tissue (5).
In certain sites, such as the liver, spleen, pituitary, adrenals, and bone marrow, the vessels that connect arterioles and venules are known as sinusoids rather than capillaries. With diameters of up to 30 to 40 m, they are generally more distended than capillaries. They have prominent fenestrations, but there are also significant gaps between endothelial cells. In the liver, there is no significant associated basement membrane.
Venules and Veins
The transition from venous capillary to muscular venule and small collecting vein is characterized by the gradual acquisition of a muscular media. Even in medium-sized veins (Figure 9.11), the internal elastic lamella is often incomplete and the muscle fibers are only poorly oriented into circular and longitudinal layers. The paracortical or high endothelial venules of lymph nodes have an important role in T-lymphocyte recirculation (32). The endothelial cells of postcapillary venules have a prominent cuboidal or columnal appearance, usually with an ovoid nucleus and a single central nucleolus. They stain specifically with the HECA-452 antibody (see below).
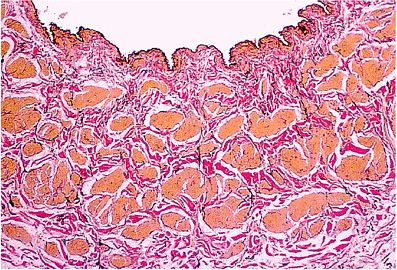 |
Figure 9.11 A renal vein from a 58-year-old woman, close to the junction with the inferior vena cava. There is no significant intimal thickening, and a thin internal elastic lamella can be identified. Note the thin layer of subendothelial collagen. The muscular wall is composed of coarse fascicles, which are not clearly arranged into circular and longitudinal layers (elastic van Gieson). |
P.226
Placental, dural, and retinal veins and the veins of erectile tissue have very little muscle. In general, the veins of the lower limb have thicker walls than those of the arm and abdomen. Most veins have valves to prevent the reflux of blood. The increasing use of the saphenous vein as an arterial conduit has led to a greater understanding of the normal structure of larger veins and the changes that occur in them as a result of aging. Large veins have a thin layer of subendothelial connective tissue with one or more incomplete elastic lamellae. Around this, there is an inner longitudinal and outer circular smooth muscle coat. The connective tissue adventitia is often well developed. Saphenous veins in middle-aged and elderly patients show intimal fibrosis and longitudinal and circular muscle hypertrophy with a substantial increase in medial connective tissue. Sometimes a prominent third outer longitudinal muscle layer (Figure 9.12) forms between the circular coat and the adventitia (33). These changes must be distinguished from the form of atherosclerosis that develops in vein bypass grafts.
Lymphatics
At the light microscopic level, small lymphatics closely resemble capillaries. In general terms, lymphatics have a larger diameter and a less regular cross-sectional profile (34). They begin as dilated channels with closed ends and anastomose freely. Although they are present in most tissues, they are rarely found in the epidermis, nails, cornea, articular cartilage, central nervous system, or bone marrow. Lymphatic channels have numerous valves and are often slightly distended at these sites, producing a slightly beaded appearance. Lymphatics with a diameter of more than 0.2 mm usually have a thin muscular media, with no clear division into circular or longitudinal coats, and a fibrous adventitia. A longitudinal muscular layer is present in the right lymphatic and thoracic ducts. As detailed and illustrated below, lymphatic vessels stain specifically with two antibodies, D2-40 and LYVE-1.
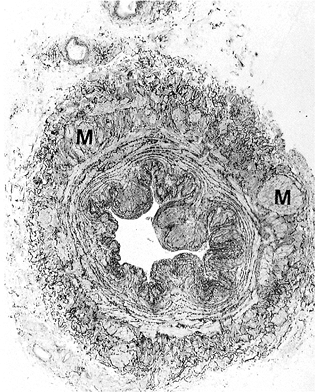 |
Figure 9.12 A saphenous vein with a prominent third outer longitudinal muscular coat (M). Reprinted with permission from: Milroy et al. Histological appearances of the long saphenous vein. J Pathol 1989;159:311 316. With permission. |
Pulmonary Arteries and Veins
Although the basic histologic structure of pulmonary vessels resembles that of their systemic counterparts, there are differences that reflect the much lower pressure of the pulmonary circuit. The lumina of major pulmonary arteries are widely dilated in comparison with wall thickness. The intima is hardly discernible. In an adult, the pulmonary arterial media is composed of only 10 to 15 parallel elastic lamellae, whereas, even in a young child, 40 aortic lamellae can be identified. The thickness of the pulmonary trunk is about 40 to 80% that of the aorta (Figure 9.13).
In the systemic circulation, the transition from elastic to muscular arteries is abrupt and is usually at the point of a major arterial orifice. In contrast, even pulmonary arteries as small as 0.5 to 1.0 mm in diameter are elastic vessels (35,36). Muscular pulmonary arteries and arterioles also have thin walls in relationship to their luminal diameter, but this may be difficult to appreciate unless special techniques of perfusion or fixation are used. In comparison with systemic arteries, there is usually a prominent internal and external elastic lamella. Arterioles give rise to a rich network of alveolar capillaries. Pericytes are not easily identified, and in places the endothelium and alveolar epithelium appear to share a common basement membrane. The walls of pulmonary veins are less structured than their systemic counterparts. The media is composed of a rather haphazardly arranged but roughly circular layer of connective tissue and muscle. No distinct and continuous elastic lamellae are present, and valves are said to be absent (Figure 9.14).
It can be difficult to distinguish the early vascular changes of pulmonary hypertension from those of normal aging (Table 9.4). The histologic changes have been described and comprehensively illustrated (35,36). The initial changes in both conditions include intimal fibrosis and medial muscular hypertrophy, and each of these features is most prominent in muscular arteries and larger arterioles (37). The absence of significant changes in the larger arteries may be misleading. In long-standing pulmonary hypertension, the complex changes in muscular arteries include florid intimal thickening, marked medial hypertrophy, and prominent dilatation of small branches of parent vessels (Figures 9.13D, 9.15). In the
P.227
most extreme examples, angiomatoid malformations may develop, and occasionally there is fibrinoid necrosis of the vessel wall (38,39). Lung biopsy is no longer used for the assessment of pulmonary hypertension in children with congenital heart disease nor in adults with primary pulmonary hypertension. However surgical pathologists must make a careful assessment of the pulmonary arteries and veins in lung biopsy specimens and be able to describe and grade these alterations accurately (40).
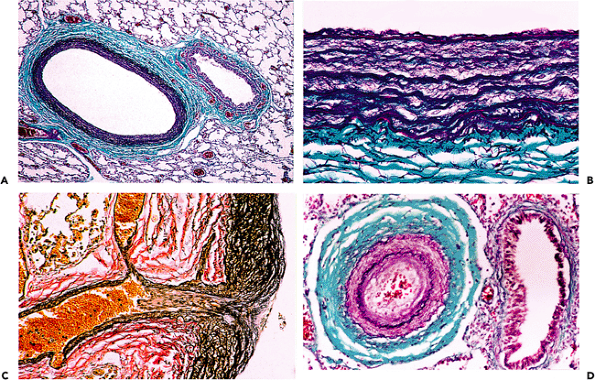 |
Figure 9.13 A. Elastic pulmonary artery from a 1-year-old child. The lung was inflated via the main pulmonary artery, which therefore appears much larger than the corresponding bronchus. B. A magnified view of the elastic wall (Gomori's trichrome). C. The transition from elastic to muscular pulmonary arteries in a 73-year-old man. Note the larger number of elastic lamellae. There is slight fibrous intimal thickening only (elastic van Gieson). D. A small pulmonary artery from a patient with longstanding pulmonary hypertension and chronic obstructive airways disease. There is hypertrophy of the muscular wall and pronounced fibrous intimal thickening (Gomori's trichrome). |
Aging changes in pulmonary veins are seldom described in detail. In severe, long-standing cardiac failure, intimal fibrosis, medial hypertrophy, and hyalinization are prominent pulmonary venous abnormalities. Marked medial hypertrophy may confer an arterialized appearance to pulmonary veins, and they may appear to have an internal and external elastic lamina. Multiple levels should be taken and stained for elastin and by a trichrome method. The elastic lamellae are seldom complete in these abnormal veins, and there is often more medial fibrosis than in corresponding pulmonary arteries. Even so, accurate distinction of abnormal pulmonary arteries and veins can be difficult.
Anastomoses, Angiodysplasias, and Vascular Malformations
There is potential for anastomoses between many arteries and veins. These are especially developed in the skin, where they contribute to thermoregulation. They vary in size from about 200 to 800 m and in some sites, such as the nail bed, have a complex structure. There are also anastomoses between pulmonary and bronchial veins and between the portal and systemic circulations. Peripheral glomus tumors almost certainly arise from supporting cells that surround the normal but rather complex anastomosing channels between digital arterioles and venules. Glomus cells do not express endothelial markers but, because they stain with smooth muscle actin and vimentin, may be related to vascular smooth muscle (41). The potential connections
P.228
P.229
between the portal and systemic circulations, either in the submucosa of the esophagus or rectum or in the periumbilical or diaphragmatic region, may be massively dilated in advanced hepatic disease. Biopsies are seldom performed surgically.
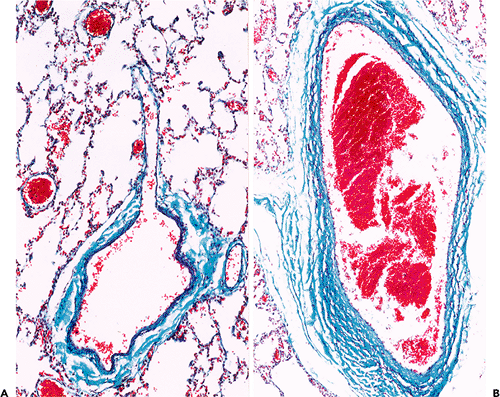 |
Figure 9.14 Normal pulmonary veins. A. A pulmonary venule draining into a small vein. Very little muscle is present in the wall. B. A large pulmonary vein close to the hilum of the lung. (Gomori's trichrome) |
Table 9.4 Histologic Features of Pulmonary Vessels | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
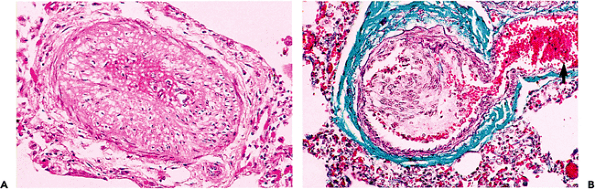 |
Figure 9.15 Advanced pulmonary hypertensive changes. A. There is marked hypertrophy of the medial muscle in a small pulmonary artery. B. An early plexiform lesion with nearby dilated thin-walled branches (arrow). (Gomori's trichrome.) |
Surgical pathologists must be familiar with the normal vascular patterns of the cerebral meninges and the colonic submucosa if cerebral arteriovenous malformations and large intestinal angiodysplasia are to be accurately assessed. Each of these areas has a rich vascular supply with numerous, sometimes thick-walled, venous channels. Malformations or angiodysplasias must only be diagnosed if there is undoubted evidence of an abnormal vessel wall. Aging changes and atherosclerosis seldom involve the smaller leptomeningeal arteries. In arteries, eccentric fibrous intimal thickening or disruption of the elastic lamellae support a diagnosis of a malformation. Veins in these malformations have irregular contours, the thickness of their muscular wall may vary markedly, and the wall can be uniformly fibrosed.
Angiodysplasia of the colon is a common cause of lower gastrointestinal hemorrhage. The lesions are usually present on the antimesenteric border of the cecum, often close to the ileocecal valve (42). They are not direct arteriovenous anastomoses but rather dilatations of preexisting, and previously normal, capillary rings and veins (Figure 9.16). The dilatation of these vessels may be the result of increased colonic muscular pressure causing intermittent obstruction of draining vessels. Multiple blocks must be examined and the appearances contrasted with a control section of submucosa from a normal colon. Submucosal arteries of the large intestine may show pronounced age-related tortuosity, and this must not be interpreted as an abnormality. A proportion of cases with good clinical or radiologic evidence of angiodysplasia will not be confirmed histologically. Some cases of massive gastrointestinal hemorrhage result from abnormally large submucosal arteries. This is most common in the stomach but also has been reported in the large and small intestine. Arteries in the submucosa of the proximal portion of the stomach can arise directly from omental vessels and may have a larger caliber than superficial arteries arising from a submucosal plexus, the so-called caliber-persistent artery or Dieulafoy's lesion (43,44).
Vascular malformations are congenital lesions composed of mature but often malformed (dysplastic) blood vessels. They result from dysregulation in the signalling pathways of vasculogenesis in early embryonic life (45) and must be distinguished from true angiomas and reactive hyperplasias. They may be solitary lesions or be part of a dysmorphic syndrome and grow slowly but progressively, usually commensurate with the growth of the patient (46,47).
P.230
They are classified according to the size of the predominate type of vessel. Clinically, a distinction is made between low- and high-flow lesions. The latter are usually arteriovenous malformations characterized by connections between feeding arteries and draining veins, without an interconnecting capillary bed, the so-called arteriovenous fistula (48). Fistulae are rarely found in tissue sections; but in these lesions arteries have a tortuous course, and a proportion of veins may show intimal thickening with collagen and elastin deposition in their walls. Pure venous malformations are composed of dilated vascular channels with walls of variable size, showing irregular degrees of attenuation and fibrosis. Complications include thrombosis with organization, papillary endothelial hyperplasia (Masson's pseudotumor), and nodular calcification. In lymphatic malformations, the vascular channels vary considerably in size and may have an incomplete muscular wall. As in other vessels their endothelium stains with CD31 and CD34 antibodies and with factor VIII-related antigen.
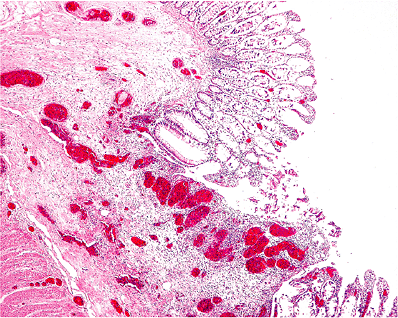 |
Figure 9.16 Angiodysplasia of the colon. Note the many dilated thin-walled blood vessels in the submucosa. Although these vessels are distended, their basic structure is unaltered. |
Vascular Surgery
The changes commonly seen in vessels after surgical procedures and interventions are summarized in Table 9.5.
Endarterectomy
Patency can be restored to a partially occluded artery by drawing out a proportion of the atherosclerotic intima. The procedure is usually applied to the carotid bifurcation, the iliac, femoral, or, occasionally, coronary arteries. Ideally, the surgeon should establish a plane between the innermost media and the intima, and the atheromatous material should be removed in its entirety. At its bifurcation the carotid artery has an elastic wall, and the material removed will include layers of elastic tissue, atheromatous debris, and thrombus. Acute postoperative thrombus formation is the most important immediate complication of the procedure. Longer term complications are recurrent thrombosis, aneurysmal dilatation, and restenosis due to fibrous intimal proliferation (49).
Bypass Grafts
The pathologic changes that occur in autologous saphenous vein bypass grafts have been described in detail (50). Care must be taken to distinguish these changes from those associated with normal aging. When subjected to arterial pressure, many vein grafts dilate and most develop some fibrous intimal thickening and medial muscular hypertrophy. In time, many develop pronounced fibrous intimal thickening with areas of lipid deposition, intramural hemorrhage, and thrombosis. These appearances closely mimic atherosclerosis and are an important cause of graft failure. In one postmortem study in which saphenous vein conduits were sampled throughout their length, more than 75% narrowing was demonstrated in 11 to 26% of the segments examined (51). Grafts can sometimes be dilated by angioplasty, but redo coronary bypass procedures are now a significant part of the work of all cardiac surgery departments.
Table 9.5 Pathologic Changes After Vascular Surgery | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
|
In cardiac surgery, coronary artery stenoses are routinely bypassed with the left or right internal mammary artery. The origin of the artery from the subclavian artery is preserved, and it is then dissected away from the chest wall. There is usually a surrounding cuff of soft tissue, but some surgeons dissect this away, producing a so-called skeletalized graft. Long-term patency rates are superior to saphenous vein grafts. The caliber of the normal internal mammary artery is similar to that of distal coronary arteries. Preexisting occlusive disease is present in fewer than 5% of patients, and only occasional grafts develop atheromatous obstructions (52). In its proximal portion, the internal mammary is an elastic
P.231
artery, but the media is muscular from about the level of the fourth rib.
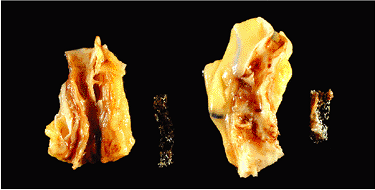 |
Figure 9.17 Coronary artery stenting. Metallic stents were inserted three weeks before this patient's death. The stent placed in the left anterior descending artery (left) is fully patent and an excellent result has been obtained by the angioplasty procedure. In contast, some reactive fibrosis has formed in the stent that was placed in the right coronary artery (right). |
Segments of the radial artery are also used as free grafts. Like saphenous vein grafts, they are anastomosed proximally to the aortic root and distally to the coronary arteries. The radial artery is muscular and is invariably free of significant atheroma.
Angioplasty
Percutaneous coronary angioplasty (PTCA) with stent emplacement is now the treatment of choice for many proximal coronary stenoses and is increasingly used as a primary intervention to open thrombosed coronary arteries after myocardial infarction (53). The mortality rate in most centers is now less than 1%, and over 90% of procedures are initially successful (52). In order to dilate the vessel, the heavily fibrous and focally calcified atheromatous plaque must be cracked open. Only when this has occurred can the deeper intima and underlying media be distended by the inflated balloon and held open by the expandable metallic stent. Early histologic studies of patients dying soon after angioplasty demonstrated a characteristic pattern of radial tears or splits, sometimes with dissections extending into the underlying media.
Stents minimize the complications of these changes, but stent thrombosis is an occasional complication (54). Restenosis is the result of fibrous intimal proliferation, thrombus formation, and an overall reduction in the size of the vessel lumen, the so-called constrictive remodelling (55,56). Stents coated with immunosuppressant or antineoplastic agents, such as sirolimus or paclitaxel, are now used routinely and have reduced rates of restenosis as compared to bare metal stent (57,58). If death occurs soon after the procedure, the stent can be carefully extracted from the opened artery (Figure 9.17), which is then processed in the usual way. After late closure, stents can be cut with an electric diamond saw and then embedded in hard plastic.
Prosthetic Vessels
Various types of fabric graft are used for the treatment of peripheral vascular disease, for closing cardiac septal defects, or in other more complex procedures in children with congenital heart disease. Acute occlusion of prosthetic vessels is usually the result of surgical technique or poor flow rates. In time, prosthetic grafts develop a pseudointima. This has a jellylike consistency, may develop a partial (though not a complete) endothelial lining, and is composed of fibrin and enmeshed leukocytes (59). However, the most striking feature of these prosthetic vessels is the intense mononuclear and giant cell reaction that develops around the woven fibers of the graft. There is usually a moderate degree of adventitial fibrosis that binds the prosthesis to the surrounding tissues and reduces its elasticity. Long-term complications include thrombosis, particularly at flexures or surgical anastomoses, infection, and deterioration of the fibers of the graft.
Electron Microscopy
Ultrastructural studies have made enormous contributions to our understanding of vascular biology. However, even surgical pathologists with a specific interest in vascular pathology have only limited experience and expertise in electron microscopy. Some of the most important ultrastructural features of vessels are summarized in Table 9.6.
Table 9.6 Ultrastructural Features of Vascular Tissues | |
|---|---|
|
P.232
Endothelial Cells
The entire vascular system is lined internally by a single layer of rather spindle-shaped endothelial cells. Small fingerlike microvilli, 200 to 400 nm long, may be seen on the surface of endothelial cells (Figure 9.18). A thin polysaccharide layer, the glycocalyx, coats the luminal surface of the endothelium. This is up to 100 nm in thickness, but its exact function is uncertain. Although endothelial cells have relatively sparse endoplasmic reticulum, a small number of free ribosomes, and an inconspicuous Golgi apparatus they produce a variety of molecules that are important in blood coagulation and the regulation of vascular tone.
Junctional complexes between endothelial cells are tight, adherens, or gap junctions (60). Tight junctions have a barrier function and help to maintain cell polarity. Molecules of the claudin family create the barrier and regulate electrical resistance between cells (61,62). Loss of this barrier function may be important in disorders such as diabetic retinopathy (63,64). Adherens junctions regulate permeability to white cells and soluble molecules and have a role in contact inhibition. Gap junctions are assembled from proteins known as connexins and form channels between adjacent cells (65). Alterations in gap junction proteins have been documented in human heart disease, including atrial and ventricular arrhythmias (66).
Inclusions of Endothelial Cells
Lysosomes are readily identified in most endothelial cells and are involved in intracytoplasmic digestion of foreign debris and products of metabolism. In many areas of the vascular system, membrane-bound vesicles measuring up to 80 to 90 nm can be identified (Figure 9.19). They are most prominent on the abluminal surface of the endothelial cell. They were originally known as plasmalemmal vesicles but are now usually termed caveolae. Their functions include the sequestration and concentration of small molecules, and they contribute to the endothelial barrier function, regulation of nitric oxide synthesis, and cholesterol metabolism (67). Weibel-Palade bodies are characteristic inclusions of endothelium and measure up to 3 m in maximum dimension. These membrane-bound structures contain up to 25 parallel tubular arrays. Immunologic studies have shown that Weibel-Palade bodies are sites of storage of von Willebrand factor. They are a useful marker of endothelial cells but are seldom as conspicuous as in Figure 9.19.
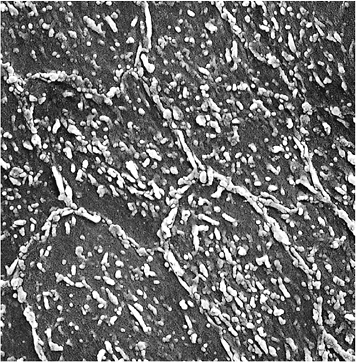 |
Figure 9.18 Scanning electron microscopic appearances of the endothelium from an experimental animal perfused under pressure with fixative. The junctions between individual endothelial cells are clearly seen; and, in this preparation, microvilli are particularly prominent (original magnification 1,200). |
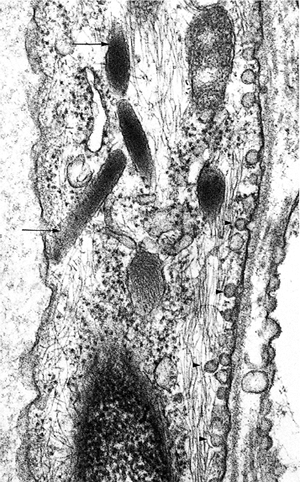 |
Figure 9.19 Transmission electron micrograph of an endothelial cell from a small subcutaneous capillary. Plasmalemmal vesicles are present on the abluminal surface (arrowheads). There are conspicuous Weibel-Palade bodies (arrows). Only part of the endothelial cell nucleus is included (bottom) (original magnification 15,000). |
The permeability of capillaries varies considerably from organ to organ. In some sites, such as the renal glomerulus,
P.233
the hepatic sinusoids, the small intestine, and some endocrine glands, there is a rapid interchange between blood and the surrounding tissue. Some of these permeability differences are related to the exact nature of the junctions between endothelial cells, but endothelial fenestrae also have an important role in this respect. These fenestrations are in fact the openings of irregular, and sometimes incomplete, transendothelial channels that allow the rapid interchange of fluid between the blood vessel lumen and the interstitium.
Media
In the human aorta, homogeneous parallel elastic lamellae alternate with layers containing smooth muscle cells and a variety of extracellular components. Smooth muscle predominates in muscular arteries. The power of contraction of smooth muscle is as great as skeletal muscle and can be maintained for longer periods with greater shortening. The structure of smooth muscle cells is maintained by the intermediate filaments vimentin and desmin, and the contractile forces are generated by actin and myosin filaments. Smooth muscle cells are arranged in parallel longitudinal bundles with the wide part of one cell opposed to the tapering part of another. Each smooth muscle cell is covered by a basal lamina which merges with fine collagen and elastin fibers (5).
Individual smooth muscle cells are often linked by communicating (gap) junctions, but tight junctions are not generally seen. In the microcirculation and in some larger arteries and arterioles, there are gap junctions between the smooth muscle cells and the overlying endothelium (68). These myoendothelial junctions could have an important role in relaying physiologic or pharmacologic stimuli between the blood vessel lumen and the media.
Adventitia and Supporting Cells
The adventitial layer consists almost entirely of collagen and elastic fibers. The thickness of this layer varies with the size of the vessel, and it may be continuous with the surrounding connective tissue. In some medium-sized veins, it is particularly well developed but in cerebral arteries may be as thin as 80 m. A layer of elastic tissue, the external elastic lamella, is present at the junction of the media and adventitia. In human material, it is seldom as pronounced as the internal elastic lamella but is prominent in many other mammalian arteries. The pericytes that are present in and among the basement membrane of capillaries and small venules superficially resemble fibroblasts. The ultrastructural appearance of their cytoplasmic filaments suggests that they are contractile, and this is further evidence that they are of mesenchymal origin (31).
Lymphatics and Veins
The smallest lymphatic vessels have wider lumina than blood capillaries and a discontinuous basement membrane. A variety of anchoring filaments bind the lymphatic endothelium to the surrounding collagenous tissues, perhaps providing the sort of support normally produced by basement membrane and enmeshed pericytes in capillaries. The ultrastructural appearances of venous capillaries, venules, and small veins mirror those seen at the light microscopic level.
Antigen Expression of Normal and Neoplastic Vascular Tissue
Endothelium
Endothelial cells cover the inner surface of the entire vascular tree, arterial, venous, capillary, and lymphatic. The most widely used antibodies are directed against von Willebrand factor (factor VIII), CD31, and CD34 (Figure 9.20). Because these antigens are present in all types of endothelial cells, they are considered to be panendothelial markers. The lectin Ulex europaeus 1 agglutinin binds to some -L-fructose containing glycocompounds and therefore to virtually all human endothelia (69). The staining pattern is sometimes more intense than with factor VIII antibodies, especially in immature vessels (70). However, all endothelial markers cross-react to some extent with other cell components. For example, in areas of hemorrhage or thrombosis, CD31 reacts strongly with platelets, macrophages, and lymphocytes, and von Willebrand factor can produce diffuse extracellular staining. Nevertheless these antibodies are indispensable for the identification of vascular tumors such as angiomas, hemangioendotheliomas, and angiosarcomas and can be helpful in the identification of tumor emboli in vascular or lymphatic channels, rather than in artifactual tissue spaces.
Antibodies that recognize proteins involved in the early steps in angiogenesis include antiendoglin (CD105) and anti-VEGF (71). In different sites in the vascular system, the endothelium may show marked heterogeneity in morphology, gene expression patterns, and related differences in functional status. However, there are only limited variations in the immunophenotypic profile of endothelial cells. Antibodies to glucose transporter protein 1 (GLUT-1 antibodies) react with the endothelium of cerebral capillaries, the placental vasculature, and one specific type of angioma the juvenile capillary angioma (Figure 9.21) (72). Another site specific antibody is anti-HECA 452, which reacts specifically with the plump endothelial cells of high endothelial venules in lymphoid tissue and postcapillary venules in chronically inflamed tissues (Figure 9.22). In inflamed tissues and in atheromatous lesions, endothelial cells undergo profound functional alterations (endothelial activation) associated with upregulation of antigens such
P.234
as ICAM-1, VCAM-1, and CD31 or with de novo expression of leukocyte adhesion molecules such as E-selectin. Until recently, there were no specific markers of lymphatic endothelium. Both D2-40 and LYVE-1 stain lymphatic endothelium specifically (Figure 9.22) (73), and D2-40 staining has confirmed the lymphatic origin of Kaposi's sarcoma (74) (Figure 9.23).
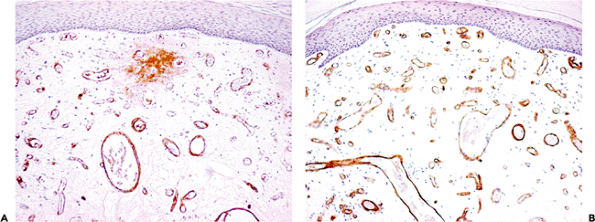 |
Figure 9.20 Staining of small vessels in a hemangioma with antibodies to factor VIII (A) and CD34 (B). As in these illustrations, the staining with CD34 is usually sharper than with factor VIII. Some nonspecific extravascular staining is often seen with factor VIII but has no significance. |
Smooth Muscle
Biochemical and immunohistologic studies have demonstrated that vascular smooth muscle has a distinctive component of contractile and intermediate filament proteins (75). In most smooth muscle, -smooth muscle actin and desmin predominate. In contrast, in vascular tissue there is abundant -smooth muscle actin, and vimentin exceeds desmin. Antibodies directed against smooth muscle actin (SMA-1) are excellent markers of medial muscle; SMA-1 recognizes the full spectrum of proliferating (or synthetic) and mature (or contractile) smooth muscle phenotypes. As SMA-1 reacts with pericytes, it clearly outlines capillaries in reactive microvascular proliferations and in pyogenic granulomas and juvenile angiomas during their growth phase. Generally all benign vascular proliferations, including glomus tumors, stain strongly with SMA-1 antibodies (Figure 9.24). In contrast, this staining is often incomplete or even
P.235
P.236
P.237
absent in angiosarcoma, hemangiopericytoma, or Kaposi's sarcoma (Figure 9.23).
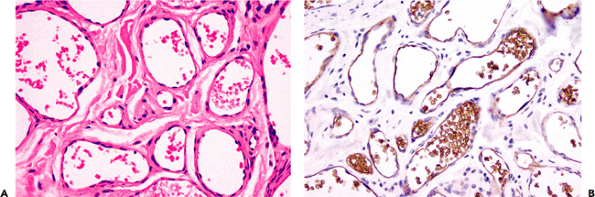 |
Figure 9.21 Site-specific staining of vascular endothelium. The vascular endothelium in this juvenile capillary hemangioma from a 3-year-old male (A) is specifically stained with the GLUT-1 antibody (B). This antibody also stains the endothelium of cerebral capillaries and the placenta. In contrast, the more commonly used endothelial antibodies such as factor VIII, CD31, and CD34 stain most types of normal and neoplastic endothelia. |
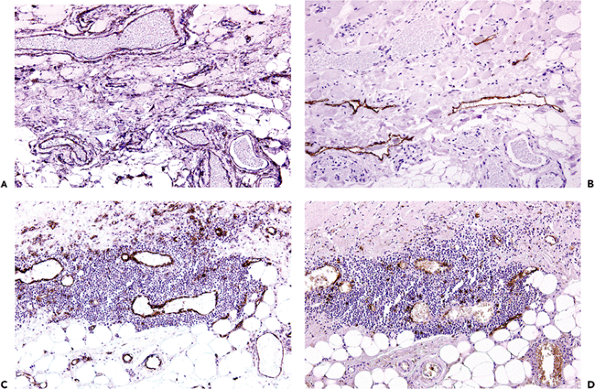 |
Figure 9.22 Immunohistochemical staining of vessels. A. and B. show a mixture of vessels from the subcutaneous tissues of a 68-year-old female from close to a leg ulcer: CD31 antibody staining identifies many vascular spaces (A), and a similar section is stained with the antibody D2-40, which recognizes lymphatic endothelium only (B). C. and D. are from a nodular inflammatory infiltrate in the aortic adventitia adjacent to a large atheromatous plaque: C has been stained with CD31, which recognizes most vessels, and D was stained with HECA-452, which recognizes high endothelial venules. |
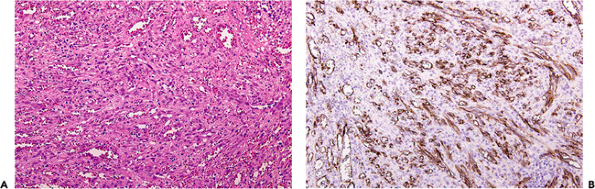 |
Figure 9.23 Kaposi's sarcoma. A. has been stained with H&E. B. was immunostained with the D2-40 antibody, a specific marker of lymphatic endothelium. Note the strong positive staining; LYVE-1 is another antibody that specifically stains lymphatic endothelium. |
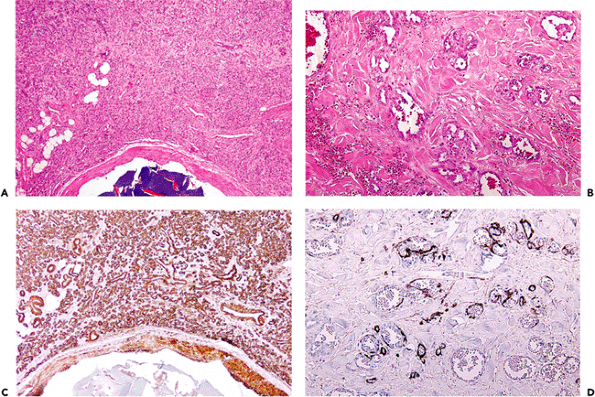 |
Figure 9.24 Patterns of staining with smooth muscle actin antibody. A. and C. are from a benign vascular proliferation. Note the intense staining of the walls of these small vessels. B. and D. are from an angiosarcoma. Only small amounts of actin are present in the walls of the malignant blood vessels (D). |
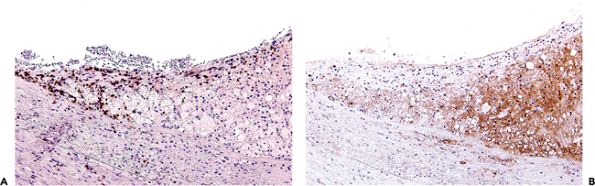 |
Figure 9.25 Immunohistochemical staining in atherosclerosis. A. CD3 positive lymphocytes are present at the edge of a lesion. B. Macrophages react for CD68. |
Other Useful Antibodies for Diagnostic Vascular Pathology
Immunohistochemical studies of the inflammatory infiltrates in atheromatous lesions (Figure 9.25) have contributed greatly to our understanding of the pathogenesis of atherosclerosis (76) but, as yet, have no value in everyday surgical pathology. T-lymphocyte markers, such as CD3 and CD4, may be of use in the diagnosis of vasculitis, especially temporal arteritis with minimal inflammatory activity (77). In transplant arteriosclerosis, there is a high relative proportion of CD8+ T lymphocytes, which also express granzyme B. In addition to von Willebrand factor (factor VIII), antifibrinogen antibodies are excellent for the demonstration of vascular leakiness and tissue damage (24,78); CD61 stains platelet aggregates in microvessels [for example in small vessel vasculitis (78)], angiolipomas, and coagulopathies [such as in the antiphospholipid syndrome]. Glycophorin A is a specific marker of erythrocytes and their precursors in the bone marrow. The epitopes are preserved in tissues for long periods, and the antibody is valuable in the detection of old hemorrhage; for example, in completely organized pulmonary thromboemboli (79) and in atherosclerotic plaques (80).
Antiamyloid antibodies (antiamyloid A, anti-immunoglobulin antibodies) are used to differentiate the nature of amyloid depositions, which have a preferential distribution in vessel walls. Cerebral vascular amyloid deposits usually do not stain with these antibodies. In pathologic conditions, such as cerebral amyloid angiopathy or amyloid found occasionally in cerebral vascular malformations, the depositions show positive staining with anti- -amyloid antibody.
References
1. Ferrari AU, Radaelli A, Centola M. Invited review: aging and the cardiovascular system. J Appl Physiol 2003;95:2591 2597.
2. Plante GE. Impact of aging on the body's vascular system. Metabolism 2003;52(suppl 2):31 35.
3. Lakatta EG, Levy D. Arterial and cardiac aging: major shareholders in cardiovascular disease enterprises: Part I: aging arteries: a set up for vascular disease. Circulation 2003;107:139 146.
4. Robins SP, Farquharson C. Connective tissue components of the blood vessel wall in health and disease. In: Sthebens WE, Lie JT, eds. Vascular Pathology. London: Chapman & Hall; 1995:89 127.
5. Wigley C. Smooth muscle and the cardiovascular and lymphatic systems. In: Stranding S, ed. Gray's Anatomy: The Anatomical Basis of Clinical Practice. 39th edition Philadelphia: Elsevier; 2005:137 156.
6. Jacob MP. Extracellular matrix remodeling and matrix metalloproteinases in the vascular wall during aging and in pathological conditions Biomed Pharmacother 2003;57:195 202.
7. Kavurma MM, Bhindi R, Lowe HC, Chesterman C, Khachigian LM. Vessel wall apoptosis and atherosclerotic plaque instability J Thromb Haemostasis 2005;3:465 472.
8. Boddaert J, Mallat Z, Fornes P, et al. Age and gender effects on apoptosis in the human coronary arterial wall. Mech Ageing Dev 2005;126:678 684.
9. Doherty TM, Detrano RC. Coronary arterial calcification as an active process: a new perspective on an old problem. Calcif Tissue Int 1994;54:224 230.
10. Elliott RJ, McGrath LT. Calcification of the human thoracic aorta during aging. Calcif Tissue Int 1994;54:268 273.
11. Maier W, Altwegg LA, Corti R, et al. Inflammatory markers at the site of ruptured plaque in acute myocardial infarction: locally increased interleukin-6 and serum amyloid A but decreased C-rective protein. Circulation 2005;111:1355 1361.
12. Mucchiano GI, Haggqvist B, Sletten K, Westermark P. Apolipoprotein A-1-derived amyloid in atherosclerotic plaques of the human aorta. J Pathol 2001;193:270 275.
13. Schlatmann TJ, Becker AE. Histologic changes in the normal aging aorta: implications for dissecting aortic aneurysm. Am J Cardiol 1977;39:13 20.
14. Hsue PY, Salinas CL, Bolger AF, Benowitz NL, Waters DD. Acute aortic dissection related to crack cocaine. Circulation 2002;105:1592 1595.
15. Ihling C, Szombathy T, Nampoothiri K, et al. Cystic medial degeneration of the aorta is associated with p53 accumulation, Bax upregulation, apoptotic cell death, and cell proliferation. Heart 1999;82:286 293.
16. Judge DP, Dietz HC. Marfan's syndrome. Lancet 2005;366:1965 1976.
17. Ghorpade A, Baxter BT. Biochemistry and molecular regulation of matrix macromolecules in abdominal aortic aneurysms. Ann N Y Acad Sci 1996;800:138 150.
18. Lopez-Candales A, Holmes DR, Liao S, Scott MJ, Wickline SA, Thompson RW. Decreased vascular smooth muscle cell density in medial degeneration of human abdominal aortic aneurysms. Am J Pathol 1997;150:993 1007.
19. Alexander JJ. The pathobiology of aortic aneurysms. J Surg Res 2004;117:163 175.
20. Ramshaw AL, Parums DV. The distribution of adhesion molecules in chronic periaortitis. Histopathology 1994;24:23 32.
21. Szyszka-Mroz J, Wozniak W. A histological study of human ductus arteriosus during the last embryonic week. Folia Morphol (Warsz) 2003;62:365 367.
22. Stary HC, Blankenhorn DH, Chandler AB, et al. A definition of the intima of human arteries and of its atherosclerosis-prone regions. A report from the Committee on Vascular Lesions of the Council on Arteriosclerosis, American Heart Association. Circulation 1992;85:391 405.
23. Farrer M, Skinner JS, Albers CJ, Alberti KG, Adams PC. Outcome after coronary artery surgery in women and men in the north of England. QJM 1997;90:203 211.
24. Kolodgie FD, Gold HK, Burke AP, et al. Intraplaque hemorrhage and progression of coronary atheroma. N Engl J Med 2003;349:2316 2325.
25. Stary HC, Chandler AB, Glagov S, et al. A definition of initial, fatty streak, and intermediate lesions of atherosclerosis. A report from the Committee on Vascular Lesions of the Council on Arteriosclerosis, American Heart Association. Arterioscler Thromb 1994;14:840 856.
26. Stary HC, Chandler AB, Dinsmore RE, et al. A definition of advanced types of atherosclerotic lesions and a histological classification of atherosclerosis. A report from the Committee on Vascular Lesions of the Council on Arteriosclerosis, American Heart Association. Arterioscler Thromb Vasc Biol 1995;15:1512 1531.
27. van der Wal AC, Becker AE. Atherosclerotic plaque rupture pathologic basis of plaque stability and instability. Cardiovasc Res 1999;41:334 344.
28. Virmani R, Kolodgie FD, Burke AP, Farb A, Schwartz SM. Lessons from sudden coronary death: a comprehensive morphological classification scheme for atherosclerotic lesions. Arterioscler Thromb Vasc Biol 2000;20:1262 1275.
29. Schwartz CJ, Mitchel JR. Cellular infiltration of the human arterial adventitia associated with atheromatous plaques. Circulation 1962;26:73 78.
P.238
30. Hammes HP. Pericytes and the pathogenesis of diabetic retinopathy. Horm Metab Res 2005;37(suppl 1):39 43.
31. Allt G, Lawrenson JG. Pericytes: cell biology and pathology. Cells Tissues Organs 2001;169:1 11.
32. Azzali G. Structure, lymphatic vascularization and lymphocyte migration in mucosa-associated lymphoid tissue. Immunol Rev 2003;195:178 189.
33. Langes K, Hort W. Intimal fibrosis (phlebosclerosis) in the saphenous vein of the lower limb: a quantitative analysis. Virchows Arch A Pathol Anat Histopathol 1992;421:127 131.
34. O'Morchoe CC, O'Morchoe PJ. Differences in lymphatic and blood capillary permeability: ultrastructural functional correlations. Lymphology 1987;20:205 209.
35. Wagenvoort CA, Mooi WJ. Biopsy Pathology of the Pulmonary Vasculature. London: Chapman & Hall; 1989.
36. Edwards WD. Pulmonary hypertension and related vascular diseases. In: Stehbens WE, Lie JT, eds. Vascular Pathology. London: Chapman & Hall; 1995:585 621.
37. Warnock ML, Kunzmann A. Changes with age in muscular pulmonary arteries. Arch Pathol Lab Med 1977;101:175 179.
38. Rubin LJ. Primary pulmonary hypertension. N Engl J Med 1997; 336:111 117.
39. Farber HW, Loscalzo J. Pulmonary arterial hypertension. N Engl J Med 2004;351:1655 1665.
40. Patchefsky AS. Nonneoplastic pulmonary disease. In: Mills SE, ed. Sternberg's Diagnostic Surgical Pathology. 4th ed. Vol 1. Philadelphia: Lippincott Williams & Wilkins; 2004:1111 1172.
41. Weiss SW, Goldblum JR. Perivascular tumors. In: Weiss SW, Goldblum JR, eds. Enzinger and Weiss's Soft Tissue Tumors. 4th ed. St. Louis: Mosby; 2001:985 1035.
42. Warkentin TE, Moore JC, Anand SS, Lonn EM, Morgan DG. Gastrointestinal bleeding, angiodysplasia, cardiovascular disease, and acquired von Willebrand syndrome. Transfus Med Rev 2003;17:272 286.
43. Veldhuyzen van Zanten SJ, Bartelsman JF, Schipper ME, Tytgat GN. Recurrent massive haematemesis from Dieulafoy vascular malformations a review of 101 cases. Gut 1986;27:213 222.
44. Sone Y, Kumada T, Toyoda H, et al. Endoscopic management and follow up of Dieulafoy lesion in the upper gastrointestinal tract. Endoscopy 2005;37:449 453.
45. Vikkula M, Boon LM, Mulliken JB. Molecular genetics of vascular malformations. Matrix Biol 2001;20:327 335.
46. Mulliken JB, Glowacki J. Hemangiomas and vascular malformations in infants and children: a classification based on endothelial characteristics. Plast Reconstr Surg 1982;69:412 422.
47. Requena L, Sangueza OP. Cutaneous vascular anomalies. Part I. Hamartomas, malformations, and dilation of preexisting vessels. J Am Acad Dermatol 1997;37:523 552.
48. Calonje E. Haemangiomas. In: Fletcher CDM, Unni KK, Mertens F, eds. World Health Organisation Classification of Tumours: Pathology and Genetics of Tumours of Soft Tissue and Bone. Lyon, France: IARC Press; 2002:156 158.
49. Clagett GP. Cerebrovascular disease. In: Harris JW, Beauchamp RD, Evers BM, Mattox KL, Townsend CM. Sabiston Textbook of Surgery. 17th ed. Philadelphia: WB Saunders; 2004:1939 1963.
50. Garratt KN, Edwards WD, Kaufmann UP, Vlietstra RE, Holmes DR Jr. Differential histopathology of primary atherosclerotic and restenotic lesions in coronary arteries and saphenous vein bypass grafts: analysis of tissue obtained from 73 patients by directional atherectomy. J Am Coll Cardiol 1991;17:442 448.
51. Kalan JM, Roberts WC. Morphologic findings in saphenous veins used as coronary arterial bypass conduits for longer than 1 year: necropsy analysis of 53 patients, 123 saphenous veins, and 1865 five-millimetre segments of veins. Am Heart J 1990;119:1164 1184.
52. Popma JJ, Kuntz RE, Baim DS. Percutaneous coronary and valvular intervention. In: Braunwald E, Zipes DP, Peter L, Bonow R, eds. Braunwald's Heart Disease. 7th ed. Philadelphia: WB Saunders; 2005:1367 1369.
53. Keeley EC, Boura JA, Grines CL. Primary angioplasty versus intravenous thrombolytic therapy for acute myocardial infarction: a quantitative review of 23 randomised trials. Lancet 2003;361:13 20.
54. Wenaweser P, Rey C, Eberli FR, et al. Stent thrombosis following bare-metal stent implantation: success of emergency percutaneous coronary intervention and predictors of adverse outcome. Eur Heart J 2005;26:1180 1187.
55. Kearney M, Pieczek A, Haley L, et al. Histopathology of in-stent restenosis in patients with peripheral artery disease. Circulation 1997;95:1998 2002.
56. Sangiorgi G, Taylor AJ, Farb A, et al. Histopathology of postpercutaneous transluminal coronary angioplasty remodeling in human coronary arteries. Am Heart J 1999;138(pt 1):681 687.
57. Windecker S, Remondino A, Eberli FR, et al. Sirolimus-eluting and paclitaxel-eluting stents for coronary revascularization. N Engl J Med 2005;353:653 662.
58. Dibra A, Kastrati A, Mehilli J, et al. Paclitaxel-eluting or sirolimus-eluting stents to prevent restenosis in diabetic patients. N Engl J Med 2005;353:663 670.
59. Clowes AW, Kirkman TR, Reidy MA. Mechanisms of arterial graft healing. Rapid transmural capillary ingrowth provides a source of intimal endothelium and smooth muscle in porous PTFE prostheses. Am J Pathol 1986;123:220 230.
60. Bazzoni G, Dejana E. Endothelial cell-to-cell junctions: molecular organization and role in vascular homeostasis. Physiol Rev 2004;84:869 901.
61. Van Itallie CM, Anderson JM. The molecular physiology of tight junction pores. Physiology (Bethesda) 2004;19:331 338.
62. Sawada N, Murata M, Kikuchi K, et al. Tight junctions and human diseases. Med Electron Microsc 2003;36:147 156.
63. Harhaj NS, Antonetti DA. Regulation of tight junctions and loss of barrier function in pathophysiology. Int J Biochem Cell Biol 2004;36:1206 1237.
64. Hsueh WA, Quinones MJ. Role of endothelial dysfunction in insulin resistance. Am J Cardiol 2003;92:10J 17J.
65. Sohl G, Willecke K. Gap junctions and the connexin protein family. Cardiovasc Res 2004;62:228 232.
67. Gratton JP, Bernatchez P, Sessa WC. Caveolae and caveolins in the cardiovascular system. Circ Res 2004;94:1408 1417.
68. Giepmans BN. Gap junctions and connexin-interacting proteins. Cardiovasc Res 2004;62:233 245.
69. Holthofer H, Virtanen I, Kariniemi AL, Hormia M, Linder E, Miettinen A. Ulex europaeus I lectin as a marker for vascular endothelium in human tissues. Lab Invest 1982;47:60 66.
70. Ordonez NG, Batsakis JG. Comparison of Ulex europaeus I lectin and factor VIII-related antigen in vascular lesions. Arch Pathol Lab Med 1984;108:129 132.
71. Dales JP, Garcia S, Carpentier S, et al. Prediction of metastasis risk (11 year follow-up) using VEGF-R1, VEGF-R2, Tie-2/Tek and CD105 expression in breast cancer (n=905). Br J Cancer 2004;90:1216 1221.
72. North PE, Waner M, Mizeracki A, Mihm MC Jr. GLUT1: a newly discovered immunohistochemical marker for juvenile hemangiomas. Hum Pathol 2000;31:11 22.
73. Jackson DG. Biology of the lymphatic marker LYVE-1 and applications in research into lymphatic trafficking and lymphangiogenesis. APMIS 2004;112:526 538.
74. Kahn HJ, Bailey D, Marks A. Monoclonal antibody D2-40, a new marker of lymphatic endothelium, reacts with Kaposi's sarcoma and a subset of angiosarcomas. Mod Pathol 2002:15:434 440.
75. Desmouliere A, Chaponnier C, Gabbiani G. Tissue repair, contraction, and the myofibroblast. Wound Repair Regen 2005;13:7 12.
76. Hansson GK. Inflammation, atherosclerosis and coronary artery disease. N Engl J Med 2005;352:1685 1695.
77. Weyand CM, Goronzy JJ. Medium- and large-vessel vasculitis. N Engl J Med 2003;349:160 169.
78. Meijer-Jorna LB, Mekkes JR, van der Wal AC. Platelet involvement in cutaneous small vessel vasculitis. J Cutan Pathol 2002;29:176 180.
79. Arbustini E, Morbini P, D'Armini AM, et al. Plaque composition in plexogenic and thromboembolic pulmonary hypertension: the critical role of thrombotic material in pultaceous core formation. Heart 2002;88:177 182.
80. Virmani R, Kolodgie FD, Burke AP, et al. Atherosclerotic plaque progression and vulnerability to rupture. Angiogenesis as a source of intraplaque hemorrhage. Arterioscler Thromb Vasc Biol 2005;25:2054 2061.
- ERP System Acquisition: A Process Model and Results From an Austrian Survey
- The Second Wave ERP Market: An Australian Viewpoint
- Healthcare Information: From Administrative to Practice Databases
- A Hybrid Clustering Technique to Improve Patient Data Quality
- Development of Interactive Web Sites to Enhance Police/Community Relations