10. Cell fate is determined by compartments that form by the blastoderm stage
29.9 TGFβ/BMPs are diffusible morphogens |
The principle of Dorsal-ventral development in flies, amphibians, and mammals is the same. On one side of the animal, neural structures (including the CNS) develop. This is the ventral side in flies, and the dorsal side in vertebrates. On the other side, mesenchymal structures develop. This is the dorsal side in flies, and the ventral side in vertebrates. The important point here is that the same relative development is seen from one side of the animal to the other, but its absolute direction is reversed between flies and vertebrates. This must mean that the Dorsal-ventral axis was inverted at some point during evolution, causing the CNS to be displaced from the ventral side to the dorsal side. This idea is supported by the fact that the same signaling pathway is initiated on the dorsal side of flies and on the ventral side of vertebrate embryos (for review see De Robertis and Sasi, 1996).
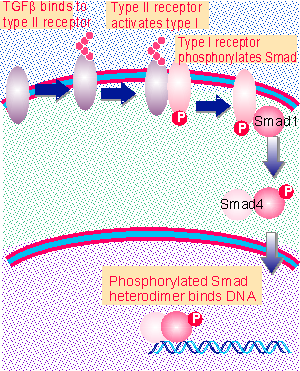 |
Figure 26.35 Activation of TGFb receptors causes phosphorylation of a Smad, which is imported into the nucleus to activate transcription. |
Mesenchymal (non-neural) structures are determined by diffusible factors in the TGFβ/BMP family. These factors are small polypeptide ligands for receptors that activate the Smads transcription factors (see Figure 26.35). Formation of neural structures requires counteracting activities that also diffuse from a center; they prevent the TGFβ/BMP ligands from activating the target receptors. (The names reflect the histories of their discoveries as transforming growth factor β and bone morphogenetic proteins; but in fact the most important role of these polypeptides is as morphogens in development.)
The involvement of this pathway in development was first described in Drosophila, where the product of dpp is a member of the TGFβ growth factor family. The receptors typically are heterodimers that form transmembrane proteins with serine/threonine kinase activity. The heterodimer consists of a type I component and a type II component. In the dpp pathway, there are two type I members (coded by thick veins and saxophone) and a single type II member (coded by punt). Mutations in tkv and punt have the same phenotype as mutations in dpp, suggesting that the tkv/punt heterodimer is the principal receptor (Ferguson and Anderson, 1992).
The activated receptor phosphorylates the product mad. This is the founding member of the Smad family. The typical pattern of activation in mammalian cells is for the regulated Smad to associate with a general partner to form a heterodimer that is imported into the nucleus, where it activates transcription.
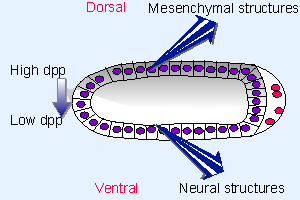 |
Figure 29.18 The morphogen dpp forms a gradient originating on the dorsal side of the fly embryo. This prevents the formation of neural structures and induces mesenchymal structures. |
Because the gene is repressed on the ventral side, dpp protein is secreted from cells only across the dorsal side of the embryo, as depicted in Figure 29.18. So Dpp is in effect the morphogen that induces synthesis of dorsal structures. Several loci influence the production of Dpp, largely by post-translational mechanisms. The net result is to increase dpp activity on the dorsal side, and to repress it on the ventral side, of the embryo. The concentration of Dpp directly affects the cell phenotype, the most dorsal phenotypes requiring the greatest concentration.
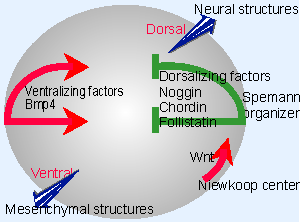 |
Figure 29.19 Two common pathways are used in early development of Xenopus. The Niewkoop center uses the Wnt pathway to induce the Spemann organizer. The organizer diffuses dorsalizing factors that counteract the effects of the ventralizing BMPs. |
The same pathway is involved in inducing the analogous structures in frog or mouse, but it is inverted with regard to the Dorsal-ventral axis. Figure 29.19 shows that Bmp4 is secreted from one side of the egg. It is antagonized by a variety of factors. Neural tissues develop in the (dorsal) regions which Bmp4 is prevented from reaching.
The crucial unifying feature is that neural tissues are induced unless the activity of dpp/Bmp4 is antagonized. Typically the Dpp/Bmp diffuses from a source, and different phenotypes may be produced by different concentrations of the morphogen. It is controversial whether the morphogen diffuses extracellularly or whether there may be a relay system that propagates it from cell to cell. Analogous pathways, triggered by different Bmps, are involved in the development of many organs.
Antagonists to Dpp/Bmp may act at the level either of sequestering the ligand itself or of acting upon its receptor. They take the form of large proteins Xchordin, noggin, follistatin in Xenopus. They induce neural tissue by inhibiting Bmp4 action. The antagonists themselves may be regulated, typically by proteolysis. Inactive Bmp/chordin complexes are cleaved by the protease Xolloid to release active Bmp.
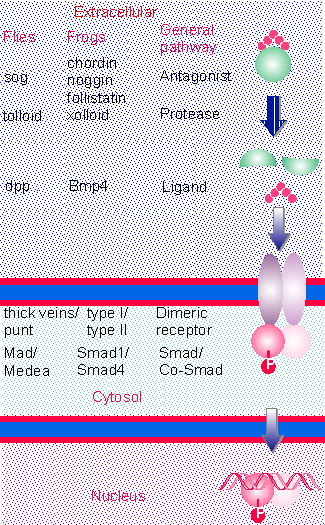 |
Figure 29.20 The TGFb/Bmp signaling pathway is conserved in evolution. The ligand may be sequestered by an antagonist, which is cleaved by a protease. Ligand binds to a dimeric receptor, causing the phosphorylation of a specific Smad, which together with a Co-Smad translocates to the nucleus. to activate gene expression. |
Figure 29.20 compares the pathways in fly and frog. The fly pathway is well characterized for dorsal-ventral development. Mutants in sog and tolloid suggest that their products antagonize and assist Dpp, respectively. The biochemical reactions actually have been better characterized for the corresponding frog proteins (Chordin is related to Sog). Frogs may have several such pathways, with a variety of Bmp ligands that interact in an overlapping manner with a family of receptors. The frog pathway shown in the figure is for the ventralizing effects of Bmp4, but the others are similar, although their specific effects on morphogenetic determination are of course different. There can be variation in specificity at each stage of the pathway. The antagonists, ligands, and receptors may be expressed in different places and times, providing specificity with regard to local concentrations of the morphogen, but there may also be partial redundancy. Both noggin and chordin must be knocked out in mouse to produce a phenotype. Each receptor has specificity for certain Smads, so that different target genes can be activated in different tissues.
| Reviews | |
| De Robertis, E. M. and Sasi, Y. (1996). A common plan for Dorsal-ventral patterning in Bilateria. Nature 380, 37-40. | |
| Research | |
| Ferguson, E. L. and Anderson, K. V. (1992). decapentaplegic acts as a morphogen to organize dorsal-ventral pattern in the Drosophila embryo. Cell 71, 451-461. | |