3. Cancer
Authors: Corwin, Elizabeth J.
Title: Handbook of Pathophysiology, 3rd Edition
Copyright 2008 Lippincott Williams & Wilkins
> Table of Contents > Unit I - Fundamental Mechanisms of Health and Disease > Chapter 3 - Cancer
Chapter 3
Cancer
Cancer is the growth of abnormal cells that tend to invade neighboring tissue and spread to distant body sites. It is a condition of uncontrolled cellular proliferation that knows no limits and serves no purpose for the host. The term cancer refers to more than 100 forms of the disease. Although each cancer has unique features, all cancers develop by following a few shared processes that in turn depend upon crucial genetic alterations. For a cell to become cancerous, these genetic alterations must: spur cell growth; inactivate genes that normally slow growth; allow cells to keep dividing, thereby immortalizing them; and allow cells to live on with abnormalities that otherwise would cause them to undergo cell suicide (apoptosis). In addition, genetic alterations must occur that allow cancer cells to recruit normal cells to support and nourish them, and to develop strategies that prevent the immune system from destroying them. These processes that cancer cells share and that differ substantially from those of normal cells are the focus not only of this chapter but of the ongoing, world-wide research effort to prevent and cure cancer.
Physiologic Concepts
Cellular Reproduction
Although all cells reproduce during embryogenesis, only certain cells continue to do so after the first few months following an infant's birth. Cells that reproduce, such as liver, skin, and gastrointestinal cells, duplicate their DNA exactly and then split into two new daughter cells. Cells reproduce by going through a process called the cell cycle, described fully in Chapter 2.
P.60
Cells that do not reproduce after birth, such as skeletal muscle cells, do not go through this cycle. Advancement through the cell cycle is tightly controlled and can be stopped or started depending on the conditions of the cell and the signals it receives, some of which are described below.
Rates of Cellular Reproduction
Cells that reproduce go through the cell cycle at some inherent rate. This rate may be increased or decreased. Cells that reproduce slowly, or not at all, spend most of their time in the gap stages (G1 or G0) of interphase. Cells that divide continually spend less time in these stages, and move frequently through the cell cycle.
Control of Cellular Reproduction
Cell cycling is controlled by the contributions of a variety of genes that respond to cues on cell crowding, tissue injury, and growth needs. In general, cells go through the cell cycle when stimulated to do so by hormones and growth factors secreted by distant cells; by locally produced growth factors; and by chemical cues released from neighboring cells, including cytokines produced by immune and inflammatory cells. These external cues act by binding to specific receptors on the plasma membrane of the target cell. Once bound, the receptor complex activates a second messenger system, which delivers the growth signal to the nucleus. When the signal reaches the nucleus, certain proteins there, called transcription factors, turn on or off specific genes that in turn produce proteins that control cell proliferation. Activated genes also produce proteins that feed back on each of the steps of signaling and messenger stimulation to amplify or minimize the effects of the initial stimulus.
The following discussion describes the external cues controlling cell growth and provides an example of an important second messenger system. Finally, the two broad categories of genes whose end products ultimately control the cell cycle are presented: the tumor suppressor genes and the proto-oncogenes.
Hormones and Growth Factors That Control Cellular Reproduction
Various hormones and growth factors may stimulate cells to increase or decrease their rate of reproduction. Epidermal growth factor (EGF), fibroblast growth factor, erythropoietin (EPO, which stimulates red blood cell proliferation), and insulin-like growth factors (which stimulate fat and connective tissue cell proliferation) can turn cellular reproduction on. Platelet-derived growth factor stimulates production of connective tissue cells. Some of these substances inhibit growth of other cells while stimulating cell division and growth in target cells.
Chemicals That Control Cellular Reproduction
Various chemicals may stimulate cells to increase or decrease their rate of reproduction. These chemicals may be released by injured or infected
P.61
neighboring cells or by immune and inflammatory cells drawn to an area after tissue injury. Many cytokines, released by cells of the immune system, stimulate cellular proliferation and differentiation. Cells possess receptors on their plasma membranes for immune and inflammatory mediators. Binding by some of these substances causes cells to produce more receptors for other immune proteins, thereby amplifying the initial response.
Physical Cues That Control Cellular Reproduction
Neighboring cells appear to communicate with each other about tissue crowding and tissue type by releasing locally active chemicals, and by passing ions and other small molecules through channels called gap junctions. Normal cells respond to physical and chemical cues put out by a large number of similar cells by slowing or stopping their rate of reproduction. This allows cellular growth and proliferation to be controlled based on tissue space requirements. These methods of communication allow cells to recognize other cells of the same type (e.g., kidney cells recognize other kidney cells).
Cytoplasmic Second Messenger System Controlling Cellular Reproduction
The cytoplasmic signal cascade begins after a protein hormone, growth factor, or other chemical binds to a cell membrane receptor and turns on a specific second messenger system. Activated second messenger proteins relay the growth-controlling signal to the nuclear transcription proteins. An example of an important cytoplasmic messenger is the ras protein. The normal ras protein transmits stimulatory signals from bound growth factor receptors on a cell's membrane to other proteins down the line that ultimately turn on cell cycling. As discussed later, many cancer cells show a mutation in the gene that produces the ras protein, such that it is always produced, even when growth factor receptors are not stimulated. This mutation results in runaway cellular proliferation. Hyperactive ras proteins are found in approximately one-fourth of all human tumors.
Tumor Suppressor Genes
Several different genes, called tumor suppressor genes, control cell cycling by coding for proteins that inhibit cellular growth and reproduction. Tumor suppressor genes are vitally important in all normally functioning cells. As described later, although cancer results from many accumulated mutations, the first mutation that sets a cell on its way to becoming cancerous often occurs in one of the tumor suppressor genes.
Tumor suppressor genes act by producing proteins that slow down or stop the second messenger brigade, including proteins that interfere with the functioning of the stimulatory ras protein. Tumor suppressor genes may also code for proteins that make up surface receptors that bind growth-inhibiting hormones or factors. Other tumor suppressor genes, when activated, stimulate a damaged cell to undergo apoptosis (programmed cell death). Finally, some tumor suppressor genes produce proteins that code for important brakes that act directly on cells about to
P.62
commit to going through the cell cycle; these genes include the RB gene and the p53 gene.
The Rb Gene
The RB gene codes for the pRB protein, the master brake of the cell cycle. Without this protein, the cell cycle is constantly in the on mode, and cellular reproduction can occur nonstop. Mutations in this gene have been identified in a variety of human cancers including bone, bladder, small cell lung, and breast cancer, and the cancer after which the gene was named, retinoblastoma.
The P53 Gene
The p53 gene codes for the p53 protein, which normally monitors the health of the cell and the integrity of cellular DNA. The p53 protein can act as a powerful brake to halt cell division before it is too late if errors in DNA transcription are present or if cellular conditions are not right. The p53 protein can cause the cell either to pause in the cycle indefinitely until a DNA error is corrected, or to undergo apoptosis. By controlling cellular replication, the p53 gene ensures that a genetic error is not passed on and only healthy cells reproduce. Mutations in the p53 gene are shown to occur in at least 50% of all types of human tumors.
Proto-oncogenes
Proto-oncogenes are genes found in all cells that, when activated, stimulate a cell to go through the cell cycle, resulting in cellular growth and proliferation. These genes may stimulate cell cycling at all levels, including (1) producing proteins that make up membrane receptors for growth-stimulating hormones and chemicals, (2) increasing the production of second messenger proteins, including the ras protein, that transfer growth signals to the nucleus, and (3) producing transcription factors that turn on vital genes that force cell growth forward (e.g., the family of myc genes).
The MYC Genes
The MYC genes are a family of proto-oncogenes that code for transcription factor proteins that drive cellular reproduction. In healthy cells, myc genes are activated only in response to growth factors acting on the cell surface. In many types of cancer, however, the myc gene is turned on constantly, even in the absence of growth factors. Cellular proliferation can occur without control when this gene is damaged.
When normal proto-oncogenes become overactive and cause uncontrolled cell division, they are called oncogenes, or cancer-causing genes.
Cellular Differentiation
Normal cells differentiate during development. Differentiation means that a given cell becomes specialized in structure and function, and aggregates with similar differentiated cells. For example, some embryonic cells are destined to become cells of the retina, whereas others are destined to become cells of
P.63
the skin or heart. The more highly differentiated a cell, the less frequently it will go through the cell cycle to reproduce and divide. Neural cells, which do not reproduce, are highly differentiated cells. Cells that seldom or never go through the cell cycle are unlikely to become cancerous, while cells that go through the cell cycle frequently are more likely to become so.
Differentiation appears to occur from selective suppression of certain genes in some cells, whereas in other cells those same genes are active. Differentiation of each cell and tissue appears to affect differentiation of neighboring cells and tissues. Cells release specific growth factors that initiate or guide differentiation of neighboring cells.
Cell Recognition and Adhesion to Like Cells
Normal cells adhere to others of the same type and group together. Although the mechanism by which cells recognize each other is not well understood, it appears to involve chemical cues secreted only by certain cells and bound by receptors present only on similar cells. Surface proteins present on one cell type that match up with proteins on similar cells, also appear to assist in cells recognizing similar cells. These surface proteins are described as adhesion molecules, and appear to hold the cells together. Cell-to-cell recognition is demonstrated by placing cells of many different types together in a Petri dish; after a certain period, the cells will have moved into clusters with only same-type cells in each cluster.
Other adhesion molecules exist between cells and the underlying tissue matrix. These connections anchor cells to one location. When normal cells become detached from each other or experience a loosening of their attachment to underlying tissue, they respond by initiating apoptosis, which prohibits cells from floating free of their tissue of origin.
The Cell Clock
Normal human cells reproduce a predictable number of times, after which they stop and become senescent. This predictability implies that cells possess some counting system that tells them when to stop dividing. This system is important because if cells divided indefinitely we would have many more cells than is compatible with life. The mechanism by which cells tick off their own divisions involves a telomere-based counting system.
Telomeres, described in Chapter 2, are the end pieces of chromosomes that shorten with each division. When the telomere length becomes sufficiently short (indicating that it has divided a certain number of times) the cell stops dividing. Putting the brakes on cell division in response to telomere shortening requires that the cell has functioning RB and p53 proteins. Occasionally, a cell continues to divide after the telomere reaches its threshold length; usually these cells soon self-destruct as their chromosomes begin to chaotically fuse and randomly break.
Cell crowding also results in neighboring cells releasing signals that inhibit the further replication of cells. This is called contact inhibition.
P.64
Pathophysiologic Concepts
Uncontrolled Cellular Reproduction
Cancer cells do not respond to the normal cues controlling cellular reproduction. Instead, they go through the cell cycle more often than normal, resulting in an overabundance of abnormal cells. Cancer cells spend little time in the gap stages of interphase and are frequently found in the M (mitosis) and S (DNA copying) stages.
Uncontrolled cellular reproduction occurs when cells become independent of normal growth control signals. This characteristic of cancer cells is called autonomy. Autonomy results when cells do not respond to the cues controlling contact inhibition for example, growth inhibitors released by neighboring cells or inhibitory growth factors and hormones traveling in the circulation. Cancer cells may disregard these signals by not producing membrane receptors that bind the inhibitory growth signals or by not activating appropriate second messengers that transmit inhibitory information to the nucleus. Other cancer cells may overproduce membrane receptors that respond to growth stimulatory signals. Cancer cells may also produce their own growth factors that bind to their cell membranes, thereby promoting self-proliferation and allowing them to be independent of any outside control.
When placed in an in vitro experimental setting, cancer cells aggressively grow on top of each other and produce layers of disorderly cells, ignoring not only chemical signals but the tendency to respect neighboring borders. Autonomy is demonstrated in the tendency of cancer cells to detach from neighboring cells and spread to distant body sites. It has been suggested that the adhesion molecules that exist between cells of the same type and between normal cells and the extracellular matrix no longer exist for cancer cells. Cancer cell autonomy may result from the inactivation of tumor suppressor genes or the change from proto-oncogenes to oncogenes.
Anaplasia
Anaplasia refers to regression of a differentiated cell to a less differentiated stage. Cancer cells demonstrate various degrees of anaplasia. By undergoing anaplasia, a cancer cell loses its ability to perform its previous functions and bears little resemblance to its tissue of origin. Highly anaplastic cells may appear embryonic and begin to express functions of a different cellular type. Some cancer cells may become ectopic sites of hormone production. For instance, antidiuretic hormone (ADH) or adrenocorticotrophic hormone (ACTH), hormones that are normally synthesized by cells of the hypothalamus and anterior pituitary, respectively, may be secreted by ectopic sites of hormone production. Lung cancers frequently become ectopic sites of hormone production.
P.65
Because the immune system poorly responds to embryonic antigens, the presence of highly anaplastic cells may interfere with the host's immune response to the tumor and usually indicates a particularly aggressive cancer.
Loss of the Cell Clock
Many cancer cells secrete an enzyme, telomerase, that acts to replace the telomere ends of chromosomes that shorten with each cell division. This leads to a destruction of the cell counting system and immortality of the cell. Not only does telomere replacement allow a cancer cell to continue to divide, increasing its number, but it also gives the cancer cell time to accumulate more mutations, some of which may improve the cell's ability to evade the immune system or produce newer, more potent growth-stimulatory factors.
Nuclear and Cytoplasmic Derangement
Cancer cells often demonstrate multiple derangements of the nucleus, cytoplasmic organelles, and cytoskeleton. The nucleus is frequently enlarged and deformed, with obvious chromosomal breaks, deletions, additions, and translocations. The rate of mitosis is usually increased. In the cytoplasm, intracellular structures show disorganization and changes in size and shape. Changes in the microtubules that support the cell and are necessary for the control of virtually all intracellular functions are especially significant. The mitochondria become disorganized and misshapen.
Tumor Cell Markers
Some cancer cells release tumor cell markers, which are specific substances secreted by a tumor into the blood, urine, or spinal fluid of an individual with a particular cancer. Tumor cell markers may be specific antigens present on the cancer cells. Some tumor antigens are similar to fetal antigens and are called oncofetal antigens ( onco refers to cancer). Because fetal antigens often do not provoke an immune response, they may mask the tumor against the host's immune system. Tumor cell markers may even include fragments of DNA that are detectable, with increasingly sensitive measurement techniques, in the circulation when produced in excess by certain tumors.
Clinical Implications of Tumor Cell Markers
Tumor cell markers are clinically important because they offer a means of identifying certain cancers as well as a cancer's progression before, during, and after treatment. For instance, if a specific tumor cell marker is identified in a patient, it suggests that cancer may exist in the person, and further diagnostic evaluation is necessary. Furthermore, in patients with a known malignancy, if after radiation or chemotherapy the tumor cell marker is not detectable, it suggests that the cancer is in remission. If,
P.66
however, the tumor cell marker fails to decrease during therapy or reappears in high concentration after therapy, the tumor is unlikely to be in remission.
Examples of Tumor Cell Markers
Examples of tumor cell markers include:
Alpha-fetoprotein for liver and yolk sac (ovarian and testicular) cancers
Carcinoembryonic antigen for colorectal cancer
Human chorionic gonadotropin (hCG) for many tumors, including choriocarcinoma (usually cancer of the uterus)
Acid phosphatase and prostate-specific antigen (PSA) for prostate cancer
Monoclonal immunoglobulin (one subtype of antibody) for multiple melanoma
CA-125, a protein released from female reproductive organs as well as from the lining of the chest and peritoneal cavities. It increases with inflamed or injured tissue and is a marker for ovarian cancer.
Although the presence of a tumor cell marker may indicate the presence or recurrence of cancer, sole reliance on the presence or absence of a cell marker is not recommended. For example, PSA is detectable in all adult men; only an unexpected rise in PSA in a given individual, or an elevation above a certain age-dependent threshold, is suggestive of disease. Likewise, pregnant women have increased hCG, and CA-125 may be increased in women for reasons other than ovarian cancer. Failure to detect a tumor cell marker does not mean that an individual is cancer-free.
Tumor Growth Rate
Each tumor grows at a certain rate dependent on characteristics of both the host and the tumor itself. Important characteristics of the host that affect a tumor's growth rate include the person's age, sex, and overall health and nutritional status. The status of the host's immune system is also important. An individual who is immunosuppressed may be unable to recognize a tumor as foreign, or may be unable to respond to a tumor that he or she recognizes. Certain hormonal states (e.g., pregnancy) may stimulate certain tumor growth rates, while stress may affect the host's ability to restrict the development or growth of a tumor.
Important characteristics of a tumor that affect its growth rate include its location in the body and its blood supply. The degree of cellular anaplasia and the presence or absence of tumor growth factors are also important characteristics. Many tumors depend on circulating or self-produced growth factors to stimulate their growth. Therefore, the tumors that grow most rapidly often populate their surface membranes with receptors for these factors. In addition, some tumor cells secrete chemicals that make
P.67
the local environment more favorable to their growth. An example is secretion of tumor angiogenesis factors, described below.
Tumor Angiogenesis Factors
Tumor angiogenesis factors are substances secreted by tumor cells that stimulate the development of new blood vessel formation. To survive, all cells require an adequate blood supply for the delivery of oxygen and nutrients and the removal of waste products. Once a group of cancer cells has grown to a certain size (approximately 1 to 2 mm in diameter), it will outgrow its original blood vessel supply and must stimulate the development of new blood vessels to grow further.
Measuring tumor angiogenesis factors in the blood or urine may allow for early diagnosis of some cancers. Even more exciting are new treatments for cancer that involve blocking the production of tumor angiogenesis factors. Experiments demonstrate that without angiogenesis, tumors soon shrink and sometimes disappear. Interventions to block tumor angiogenesis factors in humans with cancer are a promising avenue of targeted drug therapy.
Descriptions of Tumor Growth and Spread
Growth and spread of a tumor is often described clinically; some of the different terms used are listed below. Tumor treatment often depends on the grade and stage of the cancer.
Grading: An assessment of the tumor based on the degree of anaplasia it demonstrates. For example, poorly differentiated (highly anaplastic) cells are assigned a high grade.
Staging: A clinical decision concerning the size of a tumor, the degree of local invasion it has produced, and the degree to which it has spread to distant sites in a given individual.
Doubling time: An estimate of the mean amount of time required for the division of the tumor cells. Tumor cells that rapidly divide have a short doubling time.
Tumors may grow only locally, or may spread to distant sites in the process called metastasis. It is the metastasis of tumors that may ultimately lead to death of the individual.
Local Growth of a Tumor
The term cancer refers to the crab-like projections put out by a growing tumor into the local tissue. Tumors spread locally when these crab-like projections injure and kill neighboring cells. Growing tumors injure and kill neighboring cells both by compressing the cells and blocking off their blood supply. Tumor cells also appear to release chemicals or enzymes that destroy the integrity of a neighboring cell's membrane, causing the cell to
P.68
lyse and die. When neighboring cells die, the tumor can easily grow to occupy that space. As described earlier, to grow beyond a certain size, tumors must stimulate the development of their own blood supply to meet high metabolic demands.
Metastasis
Metastasis is the movement of cancer cells from one part of the body to another. Metastasis usually occurs through the spread of cancer cells from the original (primary) site in the blood or lymph to a new, secondary site. The term malignancy refers to the ability of a tumor to metastasize. See page C1 for illustrations.
The Process of Metastasis
Steps involved in the metastasis of a primary tumor to a distant site include detachment, invasion, dissemination, and seeding (Fig. 3-1).
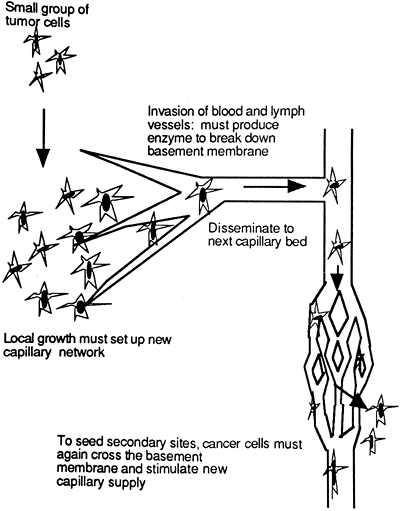 |
Figure 3-1. Tumor detachment, invasion, dissemination, and seeding. |
P.69
Detachment
To metastasize, cancer cells must first detach from their primary cluster. Recall that normal cells are linked to neighboring cells and underlying matrix tissue, and thus detach with difficulty. In addition, if a normal cell senses that it has become detached from its neighbors, it undergoes apoptosis. Cancer cells, in contrast, lose adhesion with like cells and the extracellular matrix, allowing for relatively easy detachment. Likewise, a cancer cell may produce chemicals that mimic those secreted by neighboring cells, thereby fooling its internal checkpoints into thinking that it is still attached to other cells, and thus avoiding apoptosis. Cancer cells may also deactivate tumor suppressor genes, thereby eluding apoptosis by this means as well. As cancer cells detach, they begin to invade surrounding membrane barriers and enter the circulation.
Invasion
To spread to distant sites, detached tumor cells must gain entrance to a blood or lymph vessel. To do so, the tumor cells must: (1) cross the basement membrane (a thin layer of tissue) separating its tissue of origin from the rest of the body, and (2) cross the basement membrane of a local blood or lymph vessel. To break down basement membrane walls and gain access to the circulation, tumor cells secrete specific enzymes that attack the integrity of the tissue. One such enzyme secreted preferentially by cancer cells to break down walls of a capillary is collagenase type IV. Collagenase type IV is effective in the spread of cancer cells because the new blood vessels formed in response to tumor angiogenesis factor are relatively thin and easily breached.
Dissemination and Seeding
Movement of tumor cells in the blood or lymph is called dissemination. Eventually, and especially if they are traveling in clumps, some tumor cells will get caught in a capillary or lymph network downstream from the primary site. Although many cells may die, a few tumor cells at this new site may survive and begin to seed the area. The more cells that detach from the primary tumor, the more likely it is that at least one will survive the journey and start a new tumor growth elsewhere.
When the secondary site has reached a critical size, the tumor cells will again begin to produce tumor angiogenesis factor and new blood vessel formation will be initiated to support growth of this secondary site.
Progress of a Metastasizing Tumor
Because cancer cells tend to be large, most lodge in the nearest lymph or capillary bed downstream from the primary tumor site. Because of this, the lungs, which receive systemic venous blood directly from most organs, are the most common sites of metastasis. Venous blood from the gastrointestinal (GI) tract and pancreas travels first to the liver through
P.70
the hepatic portal blood flow system, causing the liver to be the most common site of cancers from these organs. Metastasis is evaluated by observing for secondary sites in the lymph nodes nearest to the primary site, and then progressively further from this site. If exploration of the node closest to the primary site called the sentinel node is negative for tumor cells, it is likely that more distant nodes will not have been seeded either. Exceptions to this rule are tumor cells that show a striking preference to colonize certain tissues not necessarily downstream. The classic example of this preference is the tendency of prostate cancer to metastasize to bone. It has been suggested that in these cases, complementary adhesion molecules draw the tumor cells to the distant tissue. Finally, rough handling of a tumor during evaluation or surgery may cause cancer cells to break off from the primary site, thus increasing the likelihood of metastasis. Figure 3-2 identifies the typical metastasis pattern of tumors from various locations.
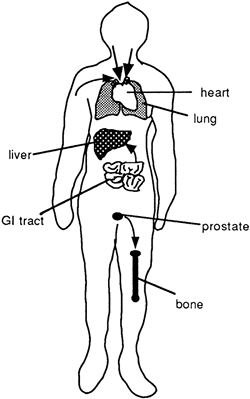 |
Figure 3-2. Spread of tumor cells from a primary site to a secondary site. Note: most tumors spread through blood or lymph to the lungs; GI tumors spread in portal flow to the liver; prostate tumors frequently spread to the bone. |
P.71
The Immune System and Cancer
The presence in the blood of antibodies, T cells, and natural killer (NK) cells produced against specific tumor antigens has been confirmed in individuals with cancer. In addition, individuals who are immunocompromised, including those with AIDS or those taking immunosuppressant drugs, have an increased chance of developing cancer. Potent anti-cancer cytokines, including tumor necrosis factor alpha (TNF ), have been identified that assist the immune system in identifying and destroying cancer cells. All of these findings demonstrate clearly that the immune and inflammatory systems have important roles in fighting and preventing cancer.
Cancer Cell Evasion of the Immune Response
Despite an apparent immune response to tumors, cancer cells are frequently able to evade the immune system. Highly anaplastic cells that primarily express oncofetal antigens are most likely to evade immune detection and are especially malignant. Other cancer cells may demonstrate changes in the expression of the major histocompatibility (MHC) antigens that normally stimulate a cell-mediated immune response, which may also allow cancer cells to evade the immune system. Cancer cells may also survive a host immune response by producing blocking antibodies that capture all host antibodies built against the tumor, allowing the tumor to continue to grow. These and other means by which tumor cells may escape immune recognition or destruction are being investigated. Experiments to boost the immune response to a tumor are also underway.
Cancer Cell Stimulation by the Immune Response
A too robust immune or inflammatory response also has been implicated in the development of cancer. For example, about 25% of cancers appear to be related to chronic infection. Liver cancer often develops after years of chronic infection by the hepatitis B or C virus, while stomach cancer is related to infection by Helicobacter pylori. Some research suggests that some of the cytokines released by white blood cells in response to infection, including TNF , may block apoptosis of damaged or mutated cells, thus leading to abnormal cell proliferation and cancer. Other pro-inflammatory cytokines may stimulate the production of angiogenesis factors normally required for wound healing or reactive oxygen species that further damage the cell.
Conditions of Disease
Cancer
There are several categories of cancer, and several theories as to how cancer develops. This section reviews the categories of cancer and presents the theory of carcinogenesis.
P.72
Categories of Cancer
Tumors are identified based on the tissue from which they develop. The suffix oma is usually added to the tissue term to identify it as a tumor, either benign or cancerous. Several general categories of cancer are presented. Individual cancers are discussed in chapters pertaining to specific organs or systems.
Carcinoma is a cancer of the epithelial tissue, including cells of the skin, testis, ovary, mucus-secreting glands, melanin-secreting cells, breast, cervix, colon, rectum, stomach, pancreas, and esophagus.
Lymphoma is a cancer of the lymphatic tissue, including the lymph capillaries, lacteals, spleen, various lymph nodes, and lymph vessels. The thymus and bone marrow may also be affected. Specific lymphomas include Hodgkin lymphoma (cancer of the lymph nodes and spleen, formerly called Hodgkin's disease) and malignant lymphoma.
Sarcoma is a cancer of the connective tissue, including cells found in the muscle and bone.
Glioma is a cancer of the glial (support) cells of the central nervous system.
Carcinoma in situ is a term used to describe abnormal epithelial cells that are as yet confined to a certain area and thus considered preinvasive lesions.
The Theory of Carcinogenesis
Cancer development is a multi-step process that usually requires decades to occur. The first step in carcinogenesis is thought to be a mutation in the DNA of an individual cell during DNA replication (copying). Although mistakes in DNA copying are not unusual, most mistakes are identified by proofreading enzymes that travel down the DNA strands to check for errors, signaling the cell cycle to stop for repair when necessary. If a mistake cannot be repaired, the cell normally is instructed to self-destruct.
The theory of carcinogenesis suggests that in certain individuals, an error in DNA copying may not be noticed, the cell cycle may not stop in time for repair, or the defective cell may not self-destruct. If the DNA error is not identified and corrected, the genetic change becomes a permanent mutation and is passed on to all daughter cells. This step is irreversible and is called cellular initiation. For a cancer to develop from this first, irreversible event, years of additional interactions with the cell by endogenous (internally-produced) and exogenous (environmental) factors that cause additional genetic changes must occur, and all must lead to the production of a cell that proliferates aggressively and without quality control. These additive effects are called promoting events. If the promoting events have significance related to a cell becoming autonomous, they may cause the cell to become cancerous (Fig. 3-3). Factors that promote the acceleration of the cell cycle through stimulation of oncogenic genes and those that allow an abnormal cell to avoid detection by the immune system are most likely to result in a mutated cell becoming carcinogenic.
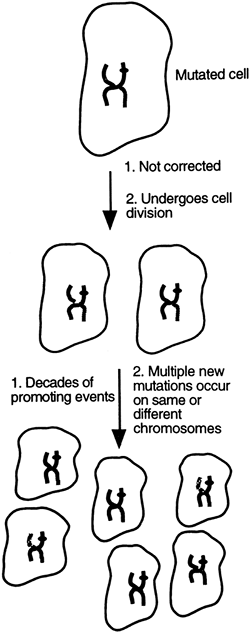 |
Figure 3-3. An uncorrected mutation passes to new daughter cells. If promoting events and new mutations occur, the cell may be released from normal growth controls to proliferate uncontrollably. |
P.73
A key point in this scenario is that the failure to detect or repair a DNA error is the first step of the cascade. This failure usually occurs in an individual who inherited a mutation in a tumor suppressor gene from one parent and developed a mutation in the other gene later in life, or in an individual who developed, over the course of a lifetime, mutations in both of the genes coding for a particular tumor suppressor, neither of which were themselves caught and repaired. This description explains how tendencies to develop cancer can be present in families, how those with such tendencies tend to develop cancers earlier in life than those without, and how cancer incidence increases with age as more and more spontaneous mutations occur. The
P.74
multi-step theory of carcinogenesis also allows for the acceptance of many causes of DNA mutation, many different variables that interact over the span of years, and roles for both heredity and environment.
Effect of Frequent Cell Cycling on Transcription Errors
The more times DNA is copied and a cell replicates, the greater the chances of an error being made, a mistake being overlooked, and a mutation being passed on. Under some circumstances, these mutations may result in cancer. Cells that reproduce and divide frequently are at greatest risk of becoming cancerous.
Monoclonal Tumor Development
When a tumor develops, it appears to do so from a mistake passed on from a single cell. This results in a monoclonal tumor from one ancestral cell. This theory is consistent with there being one mutated cell that eventually develops into a cancer.
Promoters of DNA Replication Errors
Although some mistakes in DNA replication occur randomly, certain physical agents, chemicals, and microorganisms are known to cause DNA errors. Such agents include ionizing radiation, ultraviolet radiation, components of cigarette smoke, aromatic hydrocarbons, certain dyes, nitrosamines (present in preserved meats), aflatoxin (present on moldy peanuts), and asbestos. Some of these physical agents appear to damage the DNA directly by breaking DNA base pair bonds, or indirectly by producing free radicals or other intermediates that react with and damage the DNA. Aging increases the risk of cancer in part through increased production of free radicals.
Certain viruses have been identified that can cause DNA mutations. These viruses may damage the DNA directly by causing a transcription error, or may insert into the host DNA and turn on cellular proliferation. Cancers known to be caused directly by a virus include Burkitt's lymphoma, caused by the Epstein-Barr virus, cervical cancer, caused by certain strains of the human papilloma virus, and liver cancer, caused by the hepatitis B virus. Kaposi's sarcoma also may be caused by a virus and occurs especially in those suffering from acquired immunodeficiency syndrome (AIDS).
As mentioned previously, in addition to directly altering the DNA and causing cancer, viruses and other microorganisms may irritate the cell and cause chronic inflammation. Inflammation associated with the release of pro-inflammatory cytokines may stimulate cellular proliferation and angiogenesis. Inflammation also may stimulate the cell to divide more frequently, thus increasing the likelihood of DNA transcription errors.
Effect of Any Mutating Agent
Any physical, chemical, or viral agent may cause mistakes in DNA replication or destroy proofreading enzymes. The worst-case scenario is that a mutating agent may deactivate a tumor suppressor gene that controls cell
P.75
division. If this first mistake is followed by a mutation that transforms a proto-oncogene into an oncogene, cell proliferation may occur without control. Recall from the earlier discussion that a large proportion of human cancers result from mistakes in the p53, ras, or myc family of genes and that there are two copies of each gene. It is hypothesized that cancer results from many hits: damage to one gene that is followed by subsequent damage to others. This damage requires more than one genetic error to occur in a cell over many years. Even then, cancer will not develop unless promoting effects have turned the mutation deadly.
Promotion
A mutated cell is not a cancerous cell; many years of promoting events must occur before that cell becomes cancerous. It is suggested that various promoting agents may act to cause a mutated cell to become cancerous by accelerating the proliferation of the cell. Some promoters may stimulate cellular proliferation by stimulating oncogenes or increasing surface receptors for growth factors.
Other promoting agents may not cause an actual mutation in the DNA of a gene, but rather may cause a given tumor suppressor gene to be deactivated or silenced. This type of change in the DNA is referred to as an epigenetic change in that it is separate from a change in the genome itself. Silencing of a gene usually occurs when the DNA in the area of the gene's promoter (activator) region is methylated (has a CH3 group added to it); this process leads to the promoter region being unavailable to stimulation, and hence the gene is unable to be turned on. In the case of epigenetic changes, because the DNA itself has not been altered, the effects of the promoter may be reversible if exposure to it is stopped or if an anti-methylating drug is administered.
Examples of promoters include endogenous hormones such as estrogen, certain food additives such as nitrates and salt, drugs, cigarette smoke, and alcohol; the latter two have a synergistic effect. Some substances, for example, tobacco, may act both as an initiator of a mutation and a promoter of cell proliferation.
Risk Factors
Some mistakes in DNA replication throughout a lifetime are inevitable. However, certain conditions or behaviors, known as risk factors, can increase or decrease the likelihood of a mutation arising and a mutated cell being promoted until it is cancerous.
Risk factors for cancer include exposure to any physical, chemical, or viral substance that is known to be mutagenic, and prolonged exposure to any promoter. Mutagens may be inhaled or eaten, or may act on the skin, such as in the case of ultraviolet (UV) radiation.
Behavioral Risk Factors
Certain behaviors increase the likelihood that an individual will be frequently exposed to cancer-causing stimuli. Behavioral risk factors
P.76
include cigarette smoking and diets rich in animal fat and preserved meats. Approximately a third of all cancers in the United States can be attributed to cigarette smoking, and a third to diet. Obesity also may be an independent risk factor for cancer because of the increased accumulation of fat-soluble toxins and potentially carcinogenic hormones in fatty tissue. Even a low level of alcohol consumption is linked to an increase in breast cancer, as is a sedentary lifestyle. Compared to smoking and diet, other environmental factors, including exposure to asbestos, radon, and coal tar, account for a small percentage each of cancer cases. Radiation exposure from sunlight, the primary cause of skin cancers, accounts for less than 2% of all cancers in the United States, although in other countries, including Australia, the incidence is higher.
Other behavioral risk factors include those associated with sexual behavior. A high number of sexual partners and an early onset of sexual activity increase the risk of becoming infected with the human papilloma virus (HPV), which is associated with genital neoplasms, and the AIDS virus, which is associated with Kaposi's sarcoma. Some evidence suggests that infection with the sexually transmitted virus herpes simplex 2 may also increase cervical cancer risk. Hepatitis B virus can be passed sexually and increases the risk of liver cancer.
Hormonal Risk Factors
Estrogen may act as a promoter for certain cancers, such as breast and endometrial cancer. Because estrogen levels are high in menstruating women, the risk for developing breast cancer is increased in women who started menstruating early and reached menopause late. Delayed childbearing or choosing not to bear children increases the risk of breast cancer. This increased risk appears to be related to many years of uninterrupted exposure to estrogen. Estrogen replacement therapy in postmenopausal women appears to be associated with an increase in the risk of breast cancer.
Inherited Risk Factors
A family history of cancer, especially clustered as one type, is a risk factor for developing cancer. Genetic tendencies for carcinogenesis may involve fragile or mutated tumor suppressor genes, susceptibility to certain mutagens or promoters, faulty proofreading enzymes, or a poorly functioning immune system. Inherited defects in the p53 gene and the RB gene have been documented to be associated with a high risk of cancer. Certain cancers have a higher tendency to run in families than others. For example, although most cases of colon cancer arise spontaneously, some families carry mutations that increase the risk of this disease.
Likewise, although most cases of breast cancer arise without any clear genetic link, inheritable breast cancer accounts for approximately 5% to 19% of all breast cancers. There have been two genes identified that increase a woman's chance of developing of breast cancer: BRCA1 and BRCA2. These genes are in the category of tumor suppressor genes, but their exact function in controlling cell proliferation is still unclear. Mistakes
P.77
in one or both of these genes are present in increased frequency in women with breast cancer compared to women without breast cancer, and may account for approximately half of the heritable cases of breast cancer. The presence of a mutation in either of these genes does not guarantee the development of breast cancer; however, a woman carrying a mutation in the BRCA1 gene has a 56% chance of developing breast cancer before the age of 70 and a 16.5% chance of developing ovarian cancer. Like other cancers with a genetic predisposition, familial cases of breast cancer tend to occur at a younger age.
Pediatric cancers likely have a genetic component. In children, the development of cancer is accelerated from several decades to only one or two decades. Acceleration may occur if a child inherits in the germ line (egg or sperm) one defective gene controlling a tumor suppressor or proto-oncogene product or develops such a mutation early in embryogenesis. Later, a second gene error would cause early cancer growth. Similarly, inheriting defective genes for proofreading enzymes would increase the risk of early cancer development.
It is likely that each of us has a certain genetic tendency toward developing cancer, which results in a small percentage of individuals developing cancer without exposure to known mutagens or promoters, whereas others with long-term exposures will remain cancer-free. In general, our genetic tendency toward cancer is overshadowed by risk factors we encounter in our environment and, most importantly, by the risks we take in our lifestyles and behaviors.
Factors That are Protective Against Cancer Development
Studies suggest that women who breastfeed for at least 6 consecutive months have a reduced risk of developing breast cancer. In addition, women who have had multiple pregnancies have a reduced risk of breast cancer. These findings may relate to the decreased number of menstrual periods experienced by these women. Progesterone appears to be protective against breast cancer by inhibiting the stimulatory effects of estrogen. Progesterone is high during pregnancy, which may explain why women who have had many pregnancies have a reduced risk of breast cancer.
There is also a reduced risk of breast cancer in women who exercise even moderately. This finding may be related to reduced estrogen levels or to a decrease in fat consumption and obesity.
Dietary factors are important in reducing cancer risk. Diets rich in substances known to scavenge or remove dangerous free radicals, called free-radical scavengers or antioxidants, may reduce the risk of certain cancers. These substances include vitamins A, E, and C and folic acid, all of which are prevalent in green, leafy and colorful vegetables and fruits.
Clinical Manifestations
Cancers may be diagnosed in routine examinations before any clinical manifestations appear. When clinical manifestations develop, they are
P.78
usually specific to the tumor and its site. Some general clinical manifestations that most patients with cancer demonstrate include the following:
Cachexia is a term used to describe the general wasting of fat and protein seen in many patients with cancer. Weight loss accompanies cachexia and is common in patients with cancer, many times being the presenting complaint. There appears to be a variety of causes of cachexia, including loss of appetite, poor digestion, and the increased metabolic rate of the cancer cells as they continue to go through the cell cycle and reproduce excessively. Cancer cells have high energy demands and steal nutrients needed by other cells for survival. Metabolism of food stuffs may be altered, especially if the cancer involves the liver. Cachexia also appears to be caused, at least in part, by the presence of certain cytokines produced by the immune system to fight the cancer, including tumor necrosis factor.
Anemia occurs for many different reasons and in many different types of cancers. Most individuals with metastatic cancer eventually develop anemia. It occurs early in those with cancer of the blood-forming cells of the bone marrow. This is true whether the cancer specifically affects the red or white blood cells (leukemia). Cancers that result in chronic bleeding, such as colorectal or uterine cancer, cause anemia. Platelet abnormalities are common, contributing to blood loss. Some forms of chemotherapy and radiation may depress the bone marrow, causing anemia even in patients without previous bleeding or bone marrow disease.
Fatigue frequently occurs as a result of poor nutrition, protein malnutrition, and poor oxygenation of tissues resulting from anemia. Certain cytokines produced to support the immune response against cancer also are known to cause fatigue. Growing tumors collapse the blood supply to healthy cells while stimulating their own blood supply. They take over the nutrient and oxygen supply of normal cells, causing widespread fatigue.
Diagnostic Tools
Diagnosis of cancer involves reviewing the patient's clinical presentation, gaining information on personal habits such as smoking, and investigating the patient's genetic background for cancer.
Screening tests, such as Pap smears to detect cervical cancer, mammograms to detect breast cancer, and digital examinations of the prostate coupled with a blood assay for prostate-specific antigen (PSA) to detect prostate cancer, can help to identify cancer early in its development. Testing for other cancer markers may be performed based on suspicion.
Advanced methods to diagnose and localize cancer include radiographs, CAT scans, and magnetic resonance imaging (MRI). Special bone scans may also be used.
Non-invasive diagnostic tests for oral cancer are being developed that involve testing for the presence of salivary bacteria not normally present in healthy individuals.
Cancer diagnosis is confirmed by surgically extracting a sample from a suspicious lesion, a procedure known as a biopsy, and performing a microscopic examination of the cells.
P.79
Complications
Infections are common in those with cancer. Infections develop as a result of protein malnutrition, other dietary deficiencies, and immune suppression (especially bone marrow suppression), which often accompanies conventional therapies. Hormones released in response to the long-term stress of cancer can also cause immune suppression. Complications from surgery also may result in infections in patients with cancer. Infection is a major cause of disability and death in those with cancer.
Pain may occur as a result of the invading tumor pressing on nerves or blood vessels in the area. Compression of the blood vessels can lead to tissue hypoxia, lactic acid accumulation, or cell death. Pain also occurs because the cancer cells release lytic enzymes that directly injure cells. Pain occurs as part of the immune and inflammatory reactions to the developing cancer. Fear and anxiety can worsen the pain for many patients with cancer. Most patients with advanced cancer experience pain. It is important for health care professionals to consider all treatments available to reduce pain severity and frequency in their patients.
Pain caused by compression of nerves and blood vessels occurs especially in tissues that exist in space-limiting compartments, such as bone or brain. For example, headaches are a common manifestation of advanced brain cancer, and bone pain is common with childhood hematologic cancers and bone cancer at any age. GI pain occurs when the smooth muscle of the gut is stretched.
Treatment
Several treatments for cancer are available, as outlined below and shown in Table 3-1. All have the highest rate of success with early identification of the cancer.
Surgery has long been a treatment for cancer, with the first documented account of breast removal for cancer in 200 A.D. Surgery has a better chance of curing a cancer if used on solid, well-circumscribed tumors. Tumors that have metastasized may be treated with surgery to give the patient relief from pain of the pressure of the growing tumor on surrounding nerves. Surgery is also used to debulk the tumor, which reduces tumor burden and improves the response to chemotherapy or radiotherapy.
Radiation therapy uses ionizing radiation to kill tumor cells. Radiation works on the principle that the cells most susceptible to the damaging effects of radiation are those in the S or M stages of the cell cycle. Tumor cells are most likely to be found in those stages. Unfortunately, many normal cells are also in those stages of the cell cycle at any given time, and may be killed by the therapy. In the past, it was thought that radiation killed cancer
P.80
cells only through direct damage to the DNA. However, better understanding of tumor suppressor genes has revised the consensus on how radiation kills cells. It appears that radiation kills cells primarily by altering the DNA enough that brakes on the cell cycle, especially those put on by the p53 protein and the ras protein, are activated, leading to cell suicide. Unfortunately, many times cancer cells have deactivated normal braking genes, and therefore do not undergo apoptosis when DNA damage is present. This limits the usefulness of radiation therapy. Another limitation is the scarring of normal tissue that can occur, leading to fibrosis and reduced organ function. For some cancers for example, Hodgkin lymphoma radiation may be used alone for curative purposes. Often, radiation is used in addition to surgery or to shrink the tumor, reducing tumor load.Table 3-1. Cancer Therapy, Actions, and Effects
Cancer Therapy Mode of Action Adverse Effects Surgery Reduce tumor size to alleviate pain; prevent metastasis if used early; diagnosis Pain; deformity Radiation Damage dividing cells; stimulate apoptosis; halt cell cycle Injures and leads to death of normal cells; bone marrow depression; skin desquamation Chemotherapy Multiple actions on cells to stop progression through cell cycle; may involve combination therapy and may act selectively or nonselectively Injures and kills normal cells; anorexia; nausea; bone marrow depression Immunotherapy/Biotherapy Activate host immune system to better recognize and destroy tumor cells; specifically block enzymes and growth factors required for metastasis; allow evaluation of treatment Some drugs may cause flu-like symptoms Chemotherapy uses drugs of several different classes to destroy cells in the S, M, or initial G stages of the cell cycle. Tumors grow rapidly and therefore have the most number of replicating and dividing cells and so are most susceptible to chemotherapy. However, healthy cells are also susceptible to the damaging effects of chemotherapy. Chemotherapy is frequently used in addition to surgery or radiation therapy, but may be used alone. It also may be used for palliative purposes. Chemotherapy usually causes bone marrow suppression, which in turn causes fatigue, anemia, bleeding tendencies, and an increased risk of infection.
P.81
One emerging type of adjuvant chemotherapy involves using reproductive hormone antagonists to fight reproductive cancers. The best example of this type of drug is tamoxifen, used clinically to fight estrogen-dependent breast cancers. Tamoxifen also appears to prevent the development of breast cancer in some women at high risk of the disease. Tamoxifen and similar drugs, collectively known as selective estrogen receptor modulators (SERMs), exert estrogenic effects on some tissues, for example uterine endometrial tissue, bone, and the cardiovascular system, but exert anti-estrogenic effects on the breast. This observation may allow different drugs to be tailored to the specific needs of each woman. Clinical trials to evaluate the use of tamoxifen to prevent the development of breast cancer are continuing.
Drugs have been developed that block receptors for growth factors over-expressed on certain cancer cells. The best known of this type of drug is Herceptin (trastuzumab), which binds to and blocks HER2 receptors that are over-expressed in some women with breast cancer. HER2 receptors normally bind circulating epidermal growth factor (EGF); when overabundant, the proliferative effect of EGF is excessive. By binding to the HER2 receptors, Herceptin blocks the effect of EGF. Herceptin also may act to alert the immune system to the abnormal cancer cells, thereby targeting them for destruction. In an exciting development, when patients with metastatic breast cancer positive for over-expressed HER-2 receptors were treated with Herceptin combined with chemotherapy, they demonstrated a significant decrease in the risk for breast cancer recurrence compared with patients who received the same chemotherapy without Herceptin.
Immunotherapy is a form of cancer treatment that takes advantage of the two cardinal features of the immune system: specificity and memory. Immunotherapy may be used to identify a tumor and any sites of hidden metastasis. Immunotherapy may stimulate the host's own immune system to respond more aggressively to a tumor, or tumor cells may be attacked by antibodies developed in the laboratory. Each of these options is described below.
Fluorescence-labeled antibodies: To identify a tumor, antibodies can be produced in a culture against tumor-specific antigens taken from a patient. The antibodies can then be labeled with a fluorescent isotope and injected into the patient before, or at different times during or after treatment. If the antibodies encounter their specific type of tumor cells, the antibodies will bind to the tumor cells and the resulting fluorescence can be detected, located, and measured. This use of immunofluorescent antibodies allows clinicians to identify recurrent or metastasized tumors.
Immune stimulants: Boosting the host's natural immune response to tumor cells involves activating B and T cells to notice the presence of a growing tumor. This approach has been used in skin tumors by injecting antigens capable of stimulating the immune system. Natural immunostimulants, such as interferon or some interleukins administered to
P.82
patients with certain cancers, also appear to boost a cell-mediated response to these tumors.Attacking antibodies: Antibodies produced against specific tumor antigens are being used to attack and destroy tumor cells. For example, monoclonal antibodies have been developed against malignant B cells and used in patients with lymphoma. Various other attacking antibodies also are available for use in cancer therapy.
Therapies based on the unique molecular biology of tumor cells compared to normal cells are being developed. These treatments take advantage of the recent discoveries of how cancers grow to invade local tissues and metastasize into the blood or lymph. Examples of biologic therapies being developed against tumors include drugs that specifically block tumor angiogenesis factors and enzymes such as collagenase type IV. Tumors make their own growth factors and populate their cell membranes with receptors for these factors and other substances that stimulate their growth. Drugs that block the production of tumor growth factors and the receptors for growth factors also are under development.
Gene therapy is being developed to fight cancer. In gene therapy, pieces of DNA containing special messages are delivered to cancer cells with the hope that the cancer cell will take up the DNA and begin expressing the message for which the DNA codes. Although gene therapy is far from being available clinically, early studies have provided exciting leads. The most tantalizing developments to date have been attempts to incorporate genes that inhibit angiogenesis into mice suffering from various tumors. Other ideas include infecting cancer cells with genes that code for the production of toxic chemicals that destroy the cell when produced. Another idea involves providing cancer cells with instructions to produce a therapeutic protein that would correct a faulty tumor suppressor gene or compensate for a variety of genetic errors. Other pieces of DNA may cause the cancer cell to produce a surface receptor that would bind a specific chemotherapeutic drug. Finally, the idea that it may be possible to vaccinate against a certain cancer uses gene therapy as well. In this scenario, a cancer cell may be tagged with certain genes that make it more visible to the immune system. This could improve the host immune response and lead to destruction of all cells carrying that tag. At this time, the best option would be to tag the cancer cells with genes that express certain cytokines that, when produced, alert the immune system to the cancer. This approach combines immunotherapy with gene therapy.
DNA methyl transferase inhibitors that reverse epigenetic processes associated with the methylation of tumor suppressor genes are under investigation.
The World Health Organization has outlined treatment strategies to reduce pain and has devised an analgesic ladder for pain that includes the use of non-opioid analgesia first, followed by a weak opioid and then a strong opioid. The opioid drug morphine is the drug of choice for
P.83
moderate to severe cancer pain. Other drugs, including those derived from other plant sources and some animal sources, are useful as well.Administration of recombinant erythropoietin (EPO) to treat anemia and so reduce the debilitating fatigue experienced by many patients with cancer has proven highly successful. Erythropoietin is a hormone released by the kidney in response to hypoxia, and stimulates the production of red blood cells by bone marrow. It is contraindicated for some patients.
Exercise has been shown to be effective in reducing symptoms and improving the physical and psychosocial functioning of patients with cancer.
Cancer Prevention
Cancer prevention is the ultimate goal. Although cancer will occur in some people regardless of lifestyle or personal behavior, certain types of lifestyles and behaviors increase cancer risk, while others decrease it. Cancer prevention includes the following:
Avoidance of cigarette smoking is the number one behavior that can reduce the risk of cancer, both for the individual who smokes and for family members and coworkers exposed to secondhand smoke. Children appear to be at increased risk of developing cancer after exposure to secondhand smoke even in utero. Chewable tobacco products also are associated with an increased risk of oral cancer and should be avoided. Chewing tobacco is a major health concern in adolescent medicine.
A diet rich in fruits, vegetables, and fiber and low in animal fat has been associated with a reduction in cancer in some studies.
Avoidance of sexually transmitted diseases reduces the risks of developing cancers related to infectious processes, including cervical and liver cancer.
Cancer Detection
Early detection of cancer, while not a preventive measure, can lead to containing or destroying a cancer before it has metastasized throughout the body. Early detection depends on identifying risk factors for a specific patient and using appropriate physical examination techniques. Early cancer detection tests include self breast examination and mammography, prostate examination, self testicular examination, and regular skin examination. Some screening tests, including Pap smears, tests for intestinal polyps, and biopsies of abnormal skin lesions, may allow for intervention even before dysplastic cells (cells showing early signs of abnormality) become cancerous.
Selected Bibliography
Dowell, J.E., & Minna, J.D. (2004). The impact of epidermal-growth-factor-receptor mutations in response to lung-cancer therapy. Nature Clinical Practice, Oncology 1, 2 3.
P.84
Hirsch, F.R., & Lippman, S.M. (2005). Advances in the biology of lung cancer chemoprevention. Journal of Clinical Oncology 23, 3186 3197.
Knols, R., Aaronson, N.K., Uebelhart, D., Fransen, J., & Aufdemkampe, G. (2005). Physical exercise in cancer patients during and after medical treatment: A systematic review of randomized and controlled clinical trials. Journal of Clinical Oncology 23, 3830 3842.
Lyko, F., & Brown, R. (2005). DNA methyltransferase inhibitors and the development of epigenetic cancer therapies. Journal of the National Cancer Institute 97, 1498 1506.
Mark, J. (2005). Inflammation and cancer: The link grows stronger. Science 306, 966 968.
Min, Y., & Finn, O.J. (2006). DNA vaccines for cancer too. Cancer Immunology and Immunotherapy 55, 119 130.
National Cancer Institute (2005). NCI Challenge Goal 2015. Eliminating the suffering and death due to cancer. Downloaded on October 29, 2005 from http://www.cancer.gov/.
Patel, J.D. (2005). Lung cancer in women. Journal of Clinical Oncology 23, 3212 3218.
Quigley, C. (2005). The role of opioids in cancer pain. British Medical Journal 331, 825 829.
Romond, E.H., Perez, E.A., Bryant, J., Suman, V.J., Geyer, C.E., Davidson, N.E., et al. (2005). Trastuzumab plus adjuvant chemotherapy for operable HER2-positive breast cancer. New England Journal of Medicine 353, 1673 1684.
Rubin, A.I., Chen, E.H., & Ratner, D. (2005). Basal-cell carcinoma. New England Journal of Medicine 353, 2262 2269.
Sharma, D.C. (2005). Salivary bacteria linked to oral cancers. Lancet Oncology 6, 547.
Takkouche, B., Etminan, M., & Montes-Mart'nez, A. (2005). Personal use of hair dyes and risk of cancer. A meta-analysis. Journal of the American Medical Association 293, 2516 2525.
Tomlins, S.A., Rhodes, D.R., Perner S., Dhanasekaran S. M., Mehra R., Sun, X.W., et al. (2005). Recurrent fusion of TMPRSS2 and ETS transcription factor genes in prostate cancer. Science 310, 644 648.
EAN: 2147483647
Pages: 26