3 - Breast
Editors: Mills, Stacey E.
Title: Histology for Pathologists, 3rd Edition
Copyright 2007 Lippincott Williams & Wilkins
> Table of Contents > III - Musculoskeletal System > 5 - Joints
function show_scrollbar() {}
5
Joints
Peter G. Bullough
Introduction
Bone, cartilage, ligaments, and tendons have primarily a mechanical function: providing movement, stability, and protection. Unlike the liver or kidneys, which are composed mainly of cellular elements with a metabolic function, the connective tissues are mostly formed of an extracellular material (or matrix) made up of substances to resist the tensile and compressive forces to which they are subjected.
The Normal Joint
The ends of contiguous bones together with their soft tissue components, including cartilage, ligaments, and synovium, constitute a functioning unit: the joint. There are three types of joints. The most common is the diarthrodial joint, which is a cavitated movable connecting unit between two bones. (Hyalin cartilage covers the articulating surfaces of the diarthrodial joints, with the exception of the sternoclavicular and temporomandibular joints, which are covered by fibrocartilage.) The second type is the amphiarthrodial joint, typified by the intervertebral disk and characterized by limited mobility. The third type is the fibrous synarthrosis, such as the skull sutures, which are nonmovable joints and will not be discussed further.
Diarthrodial Joint
Histology makes more sense when we have an understanding of both the function and dysfunction (i.e., the pathology) of the joint. In the diarthrodial joint normal function is characterized by: the maintenance of stability during use; freedom of the opposed articular surfaces to move painlessly over each other within the required range of motion; and an equitable distribution of load across joint tissues. Conversely clinical joint dysfunction is characterized by instability, loss of motion, maldistribution of load, and pain.
The three interdependent aspects of normal joint function depend on the shape of the joint, the mechanical properties of the extracellular matrices of the various tissues, and the integrity of the neuromuscular control.
The Shape
Perhaps the most obvious feature of any joint is the shape of its articulating surfaces. In general, one surface is convex and the other concave. The convex side of the articulation
P.98
usually has a larger surface than the concave side. These complementary shapes permit the normal range of motion, as well as providing stability and equitable loading during use.
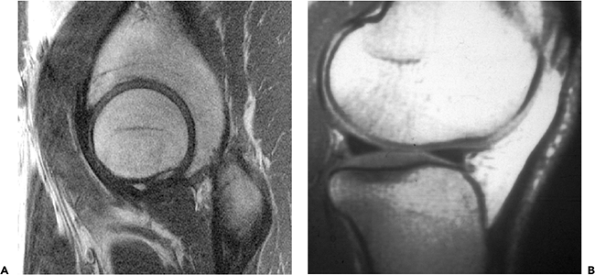 |
Figure 5.1 A. Sagittal section through the hip joint seen by MRI shows a close fit between the acetabulum and femoral head. B. Lateral MRI of a normal knee shows the gross incongruity of the articular surfaces. This is partially corrected by the interposed menisci, which act as load-bearing structures. |
In some joints at first sight the articular surfaces appear to fit exactly (i.e., they appear congruent) (e.g., the hip and the ankle) (1). However, in other joints (e.g., the knee and finger joints), it is readily apparent that the surfaces are incongruent (Figure 5.1).
In a movable joint, congruence in all positions of the joint would necessitate that all joint surfaces were either perfectly spherical or cylindrical, which obviously they are not. Therefore no joint can be congruent in all positions (though in every joint there is usually a position in which it is most congruent) (2).
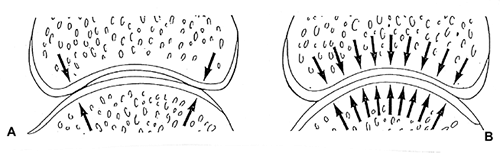 |
Figure 5.2 A. Light load. At rest and under light load, limited contact at the joint periphery assures access for synovial fluid to the joint space. B. Increased load. With increasing load, deformation of the bone and cartilage allow increased contact of the cartilage surfaces and an equitable distribution of the load. Cyclical loading permits circulation of the synovial fluid in the joint and between the articular surfaces to provide for the metabolic needs of the cartilage. |
In some joints, of which the knee is a notable example, the gross incongruencies of the opposed surfaces are partially compensated for by the interposed, pliable intra-articular fibrocartilaginous menisci (3). The menisci constitute an important component contributing to joint morphology and function and cannot be removed or damaged without significant consequences.
In many joints, perhaps most, the initial contact between the opposed articular surfaces seems to be at the periphery of the joint. However, because the tissues that make up the articulating surfaces (particularly the cartilage but also the bone) undergo elastic deformation when loaded, the surfaces come into increasing contact as the load increases, thereby distributing the load more equitably (Figure 5.2).
P.99
The incongruence, the deformation of the joint space under load, and the movement of the joint provide for the circulation and mixing of the synovial fluid, which, because the articular cartilage has no blood supply, are essential to the metabolism of the chondrocytes.
The Mechanical Properties of the Extracellular Matrices
In 1743, William Hunter (4) noted that:
The articulating cartilages are most happily contrived to all purposes of motion in those parts. By their uniform surface, they move upon one another with ease; by their soft, smooth and slippery surface, mutual abrasion is prevented; by their flexibility, the contiguous surfaces are constantly adapted to each other and the friction diffused equally over the whole; by their elasticity, the violence of any shock, which may happen in running, jumping, etc. is broken and gradually spent; which must have been extremely pernicious, if the hard surfaces of bones had been immediately contiguous.
These mechanical properties of articular cartilage (as of all other connective tissues) are determined by the extracellular matrices.
In each of the different connective tissues (bone, cartilage, ligament, etc.), as well as in each particular anatomic structure, the matrices have a unique composition and structural organization that provide for mechanical function at that locus. Disturbances in the structure and/or composition of the extracellular matrix of articular cartilage may result in joint dysfunction, and we should add that, since the joint also includes the bone beneath the cartilage as well as the capsule and ligaments, alterations in the mechanical properties of bone or disruption of the ligaments could have equally disastrous effects on joint function. Clearly some knowledge of the matrix components is necessary to an understanding of connective tissue diseases.
The connective tissue matrices are mostly synthesized and to some extent broken down by their intrinsic cells (e.g., fibroblasts, osteoblasts, osteoclasts, chondrocytes). In maintaining the physicochemical and mechanical properties of tissues, the metabolism of these cells must be subject to highly sensitive feedback systems involving both local and systemic factors.
Collagen fibers, the principal extracellular component of connective tissues, are made up of bundles of fibrils, which in turn, are composed of stacked molecules formed from polypeptide chains arranged in a helical pattern. Fourteen different types of collagen molecule are now known, and these vary both in size and configuration (5).
Type I collagen is both the most common form of collagen and the major form of collagen found in skin, fascia, tendon, ligaments, and bone. Many other types of collagen are also aligned in a staggered array to form collagen fibrils. However there are also nonfiber-forming collagens that have varying functions, such as acting as binding sites for other matrix components (type IX collagen) or facilitating calcification (type X collagen) (6).
Articular hyalin cartilage has a unique type of collagen, type II, which is structurally characterized by three triple helical -1 (II) chains. The type II fibrillar network gives articular cartilage its tensile strength and, together with the proteoglycans, is essential for maintaining the tissues volume and shape (7).
The fibrillar collagens provide tensile strength. However, connective tissues are also subjected to compression. In bone, the compressive load is resisted by hydroxyapatite. In cartilage, it is the filler between the collagen fibers that provides the compressive strength of the tissue, as well as its viscoelastic properties. This filler is composed of large negatively charged macromolecular proteoglycan aggregates.
Proteoglycans (PGs) are a group of heterogenous molecules, consisting of protein chains and attached carbohydrates, which have a sticky gel-like quality. The major PG in cartilage is aggrecan (8), containing a protein core of Mr 2.15 105 to which carbohydrate side chains (keratan and chondroitin sulfate) are attached. The core protein, which contains three globular domains, interacts with hyaluronic acid, and this interaction is stabilized by link protein. As many as 200 aggrecan molecules bind to one hyaluronic acid chain (Mr 1 2 106) to form an aggregate (Mr 5 107 to 5 108).
The highly charged PG molecules attract water and swell considerably. However, within the cartilage, the expansion of the PGs is restricted by the collagen network to approximately 20% of the maximum possible; this creates a swelling pressure within cartilage tissue. When cartilage is loaded, some water is extruded and PGs are further compressed. Removal of the load permits the imbibing of water into the tissue, together with essential nutrients, until the swelling pressure of the PGs is again balanced by the tensile resistance of the collagen network.
Aggrecan shows an age-related decrease in size and enrichment in keratan sulfate relative to chondroitin sulfate, and these changes may relate to the observed age-dependent change in the stiffness and water content of the cartilage (9).
In addition to aggrecan, the extracellular matrix of cartilage contains many noncollagenous proteins and proteoglycans, whose precise functions are only just beginning to be understood. These molecules may serve a structural or regulatory role and in some cases may do both because degradation products of some of the structural molecules are known to influence the chondrocyte. The recognition of genetic disorders in which synthesis of the matrix molecules is perturbed has aided greatly in our understanding of their functional role, but the reason for many site- and age-related restrictions in expression remains unclear. This is an area where there is still a wealth of information to be mined (10).
In a mature joint, cyclic hydrostatic fluid pressure through the entire cartilage thickness is comparable in magnitude to the applied joint pressure. Prolonged
P.100
physical activity can cause the total cartilage thickness to decrease about 5%, although the consolidation strains vary tremendously in the superficial, transitional, and radial zones. The superficial zone can experience significant fluid exudation and consolidation (compressing strains) in the range of 60%, while the radial zone experiences relatively little fluid flow and consolidation. The topological variation in the microscopic appearance and quantitative biochemistry of articular cartilage is influenced by the local mechanical loading of chondrocytes in the different zones. Patterns of stress, strain, and fluid flow created in the joint result in spatial and temporal changes in the rates of synthesis and degradation of matrix proteins. When viewed over the course of a lifetime, even subtle differences in these cellular processes may be expected to affect the micro- and macromorphology of articular cartilage.
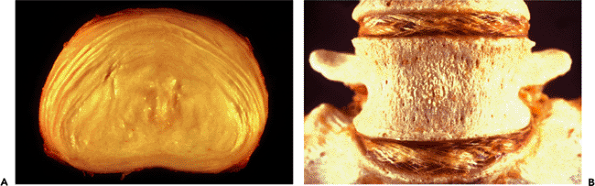 |
Figure 5.3 A. Photograph of an intervertebral disk seen in cross section. Note the layers of circumferential fibers that make up the annulus fibrosus and the well demarcated bulging central mass of the nucleus pulposus. Note also the decreasing width of the annulus from anterior (top) to posterior (bottom). B. In this desiccated specimen of the lower lumbar spine, the alternating oblique orientation of the collagen fibers in the annulus can be appreciated. |
Capsular, Pericapsular Tissues, and Muscular Control
Any consideration of functional joint anatomy must include the capsule of the joint with its synovial lining; the ligamentous conjoining of the articulating surfaces; and the neuromuscular control of joint motion. Through the perception of touch, temperature, pain, and position, sensory feedbacks monitor our movements. Correct joint function is thus dependent on intact ligaments, muscles, and nerves. As recognized by Charcot a breakdown of neuromuscular coordination can lead to profound arthritis (11).
Amphiarthrodial Joint
The intervertebral disk can be divided into two components: the outermost fibrous ring (annulus fibrosus) and the innermost gelatinous core (nucleus pulposus).
The annulus, when viewed from above, is seen to contain fibrous tissue layers arranged in concentric circles. Each layer extends obliquely from vertebral body to vertebral body, with the fibers of one layer running in a direction opposite to that of the adjacent layer. This arrangement of alternating oblique layers provides for motion that is universal in direction (rotation, flexion and extension but restricted in degree (Figure 5.3).
The fibers of the annulus are attached by Sharpey's fibers to the bony endplates of the adjacent vertebral bodies. The fibrous lamellae are stronger and more numerous in the anterior and lateral aspects of the disk than in the posterior aspect, where they are sparser and thinner. The anterior annulus is therefore almost twice the thickness of the posterior annulus. This variation probably reflects the additional protection offered by the posterior elements of the vertebral bodies. As a result of the variation in thickness of the annulus, the nucleus pulposus typically occupies an
P.101
eccentric position within the disk space, being closer to the posterior margin.
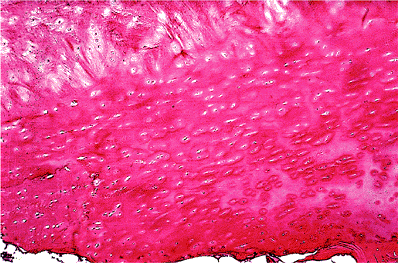 |
Figure 5.4 The nucleus pulposus of the disk (top) is separated from the bone (bottom) by a dense layer of hyaline cartilage, as demonstrated in this photomicrograph (H&E stain, 4 objective). |
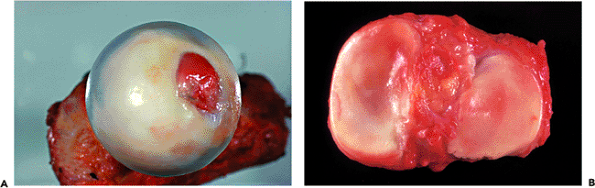 |
Figure 5.5 A. A femoral head, resected from a 16-year-old, demonstrates the blue-white translucency of young healthy cartilage. B. For comparison, the tibial plateau of a 50-year-old. The cartilage is smooth and healthy in appearance but is more yellowish in color and opaque in quality than that of the 16-year-old. |
The tissue of the nucleus is separated from that of the bone above and below by a clearly defined layer of hyalincartilage which extends to the inner margins of the insertion of the annulus (Figure 5.4).
On microscopic examination, the nucleus pulposus shows a varying number of stellate and fusiform cells suspended in a loose fibromyxoid matrix rich in proteoglycans.
Because no blood vessels are present in most of the adult disk tissue, nutrients must travel by diffusion from capillary beds at the disk margins. A restricted flow of nutrients to the nucleus and inner annulus may contribute to disk degeneration in the adult.
It should be noted that disk height, in general, is not the same in all segments of the spine, the cervical and thoracic disks being flatter than those of the lumbar region. There is also a variation in disk height from front to back, relative to the curvature of the spine. With age, the disk gets thinner as a result of age-related dehydration of the disk.
The Normal Joint Tissues
Articular Cartilage
Morphology
The articular ends of the bones are covered by hyalin cartilage, which is a nerveless, bloodless, firm, and yet pliable tissue. Hyalin cartilage deforms under pressure but recovers its original shape on removal of pressure (12). In growing children, cartilage is the most obvious precursor of the bony skeleton, and it is the means by which the bones increase in length by the mechanism of endochondral ossification through the medium of the cartilaginous growth plate (physis).
In young people, hyalin cartilage is translucent and bluish-white; in older individuals, it is opaque and slightly yellowish (13) (Figure 5.5). This change in the appearance of the articular cartilage with advancing age is also seen in other connective tissues and is probably related to a number of factors, including dehydration of the tissues, increased numbers of cross linkages in the collagen, and the possible accumulation of pigment.
On microscopic examination, articular cartilage is characterized by its abundant glassy (hyalin) extracellular matrix with isolated, relatively sparse cells located in well-defined spaces (lacunae) (Figure 5.6). It is usually described as having four layers (or zones): superficial,
P.102
intermediate, deep, and calcified. In the superficial layer, the cells are flat. In the intermediate zone, the cells have a tendency to form radial groups that apparently follow the pattern of collagen disposition. In the deep zone, the cells are hypertrophied; and in the calcified zone (i.e., the zone adjacent to the bone), the cells are nonviable and the matrix is heavily calcified (Figures 5.7 and 5.8).
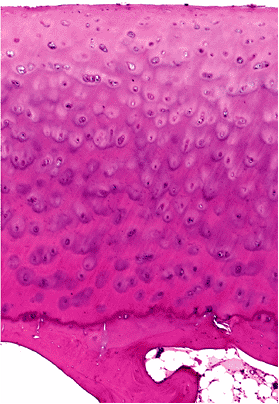 |
Figure 5.6 Photomicrograph of normal articular cartilage obtained from the femoral condyle of a middle-aged individual (H&E stain, 4 objective). |
Within the mineralized bone matrix, the cells are connected with one another by means of cytoplasmic processes; however, no such syncytial arrangement is present within the cartilage. The chondrocytes are dependent upon the diffusion of solutes through the extracellular matrix for their metabolism. Since the matrix of the deep calcified zone of the articular cartilage effectively blocks the passage of solutes from the subchondral bone, the articular cartilage is thus dependent on the diffusion of nutrients and the exchange of metabolites from the synovial fluid through the articular surface (14).
In the late nineteenth century, Hultkranz (15) demonstrated that the precisely organized fibrous system within normal articular cartilage is readily demonstrable by the simple expedient of pricking the articular surface with a pin. When this is done, a split results; if the pricking is repeated all over the surface, a pattern of split lines is revealed that is constant for each joint from individual to individual (16) (Figure 5.9).
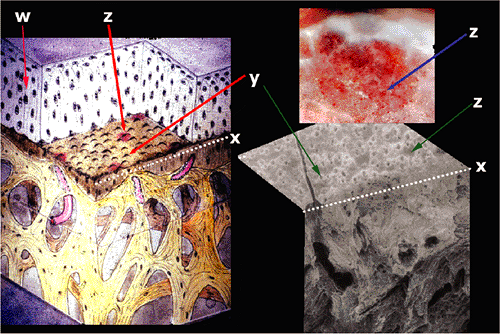 |
Figure 5.7 In this diagram of the articular surface, the organization of the articular surface seen on Figure 5.6 is shown diagrammatically. This distribution of the collagen arcades described by Benninghoff has been drawn in (w). Noncalcified articular cartilage has been removed to reveal the surface of the tidemark (x). The small volcanic structures (y) represent the location of the cells in the calcification front. The vessels that penetrate the calcification cartilage (z) are seen just beneath the calcification front. A dissection (upper right) shows the vessels when the cartilage has been pared away, and a scanning electron photomicrograph (lower right) shows the section from which the diagram was reconstructed. |
When the superficial layer of the cartilage is pared away and the exposed surface pricked, only small round holes appear instead of fissures. If the cut edge of the cartilage is pricked, a vertical split line is produced, and this occurs in all planes of section (Figure 5.10). If the fissures reflect the internal fiber arrangement of the cartilage, then at the articular surface the fibers run parallel to the surface and in the general direction of the split line, and in the deeper layers of the cartilage the fibers are predominantly vertical (17).
A combination of polarizing microscopy, transmission electron microscopy, and scanning electron microscopy have confirmed that the principal orientation of collagen fibers in articular cartilage is vertical through most of its thickness and horizontal at the surface (17) (Figure 5.11).
Electron microscopic studies have shown that, in the surface layer of normal articular cartilage, the collagen fibers are closely packed, of fine diameter, and mostly oriented parallel to the joint surface. The collagen content of cartilage progressively diminishes from the superficial to
P.103
the deep layer. In deep layers, collagen fibers are more widely separated, thicker in diameter, and are vertically aligned in such a fashion as to form a web of arch-shaped structures (18) (Figure 5.12). The collagen fibers are continuous with those in the calcified layer of cartilage but not with underlying subchondral bone. The morphology of the collagen-fibril network influences the local stresses and strains in the articular cartilage (19).
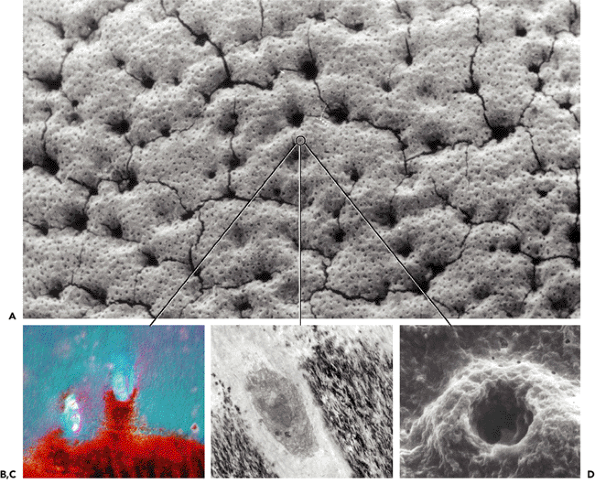 |
Figure 5.8 A. Scanning electron photomicrograph of the surface of the tidemark after the uncalcified articular cartilage had been digested away. The small dots represent chondrocytes embedded in the mineralization front (tidemark). The larger voids result from underlying vessels close to the tidemark. The cracks are preparation artifact. B. The appearance of chondrocytes embedded in this surface is shown in a cross sectional image of an H&E section, photomicrographed using polarized light. The same sample (as in B) is shown in C. as in a transmission electron and in D. as a higher power scanning image. It is hypothesized that this layer of embedded chondrocytes regulates the rate of active calcification at the tidemark. |
The menisci of the knee are composed mainly of collagen, although some PG is also present. Microscopic examination of carefully oriented sections has shown that the principal orientation of the collagen fibers in the menisci is circumferential to withstand the circumferential tension within the meniscus during normal loading of the knee joint. The few small radially disposed fibers probably act as ties to resist any longitudinal splitting of the menisci that might result from undue compression (20) (Figure 5.13).
The precise organization of collagen in the cartilage, in the annulus of the intervertebral disc, and in the menisci is also present in all the connective tissues of the body (e.g., Langer's lines in the skin) and serves a mechanical function in all of these locations.
P.104
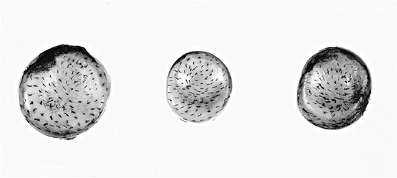 |
Figure 5.9 Photograph of the articular surfaces of three radial heads from three different individuals after the surfaces have been pricked with a pin whose tip had been dipped in India ink. Note the resulting pattern of split lines, which is unique for each joint in the body. Note that the pattern is similar from individual to individal. |
 |
Figure 5.10 Photograph of a portion of articular cartilage that has been sectioned vertically to show the cut edge and the underlying bone. The direction of pin pricks made on the surface can be seen; additional pin pricks have been made on the cut edge, all of which resulted in vertical splits. |
 |
Figure 5.11 In this polarized-light photomicrograph, the surface collagen fibers can be visualized as blue, the deeper collagen fibers (which are perpendicular) as yellow. Collagen cannot be seen in the intermediate area because the fibers in this zone are decussating as in the model of Benninghoff's arcade shown in Figure 5.7 ( 4 objective). |
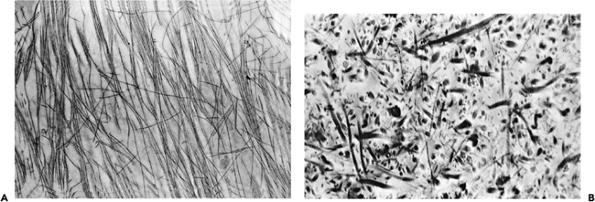 |
Figure 5.12 A. A transmission electron micrograph taken from tissue cut parallel to the surface collagen layer demonstrates thin closely packed and oriented fibers ( 10,000). B. Transmission electron micrograph, taken from tissue obtained from the midzone of the cartilage and cut randomly, demonstrates variable fiber thickness and more widely separated fibers than are seen at the surface ( 10,000). |
P.105
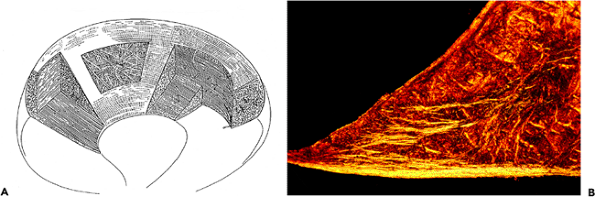 |
Figure 5.13 A. A drawing to illustrate the distribution of collagen fibers in the meniscus. The majority of the fibers are circumferentially distributed to resist the tension generated in the meniscus when the knee is under compressive load. The radially distributed fibers are most obvious on the tibial surface of the meniscus. (Modified from: Bullough PG, Munuera L, Murphy J, Weinstein AM. The strength of the menisci of the knee as it relates to their fine structure. J Bone Joint Surg Br 1970;52:564 567 with permission). B. Photomicrograph of a cross section of meniscus seen with polarized light. The tibial surface is the bottom edge where most of the fibers are radially arranged ( 1 objective). |
The distribution of PGs in the cartilage matrix is also related to the mechanical requirements. It varies quantitatively and possibly qualitatively from joint to joint, geographically within a single articular surface, and also as a function of age. (In general, PG distribution is more even in children than in adults.) The surface layers of the cartilage contain much less PG than do the deeper layers. In the deeper layers, there is a higher concentration of staining with safranin O and methylene blue around the cells (the pericellular matrix) than between the cells (the intercellular matrix) (21) (Figure 5.14).
Besides the PG aggregates, the articular cartilage contains other extracellular matrix proteins. Chondrocalcin (type X collagen) is a protein probably involved in the calcification process. Anchorin (type IX collagen) is a protein on the surface of chondrocytes involved in binding of these cells to extracellular matrix components, possibly transmitting altered stress in type II fibers to chondrocytes. Fibronectin, thrombomodulin, and cartilage oligomeric high Mr matrix protein are all found in cartilage, but their precise functions are not yet established.
 |
Figure 5.14 Portion of cartilage showing intense metachromasia around the chondrocytes in the deep part of the noncalcified cartilage. This represents staining of the proteoglycan. There is much less staining in the interterritorial matrix than around the cell. Even less staining is seen in the calcified cartilage (methylene blue stain, 25 objective). |
In histologic sections stained with hematoxylin and eosin, the junction between the calcified cartilage and the noncalcified cartilage is marked by a basophilic line known as the tidemark (Figure 5.15). This basophilic line is not seen in the developing skeleton but is clearly visible in the
P.106
adult. In older individuals (over 60), replication of the tidemark is usually evident, and in osteoarthritic joints replication may be marked (Figure 5.16). Mechanical failure in the deep cartilage rarely, if ever, gives rise to separation at the bone cartilage interface. However, when failure occurs, it is often seen as a horizontal cleft at the tidemark, presumably because of the considerable change in the rigidity of the cartilage at this junction.
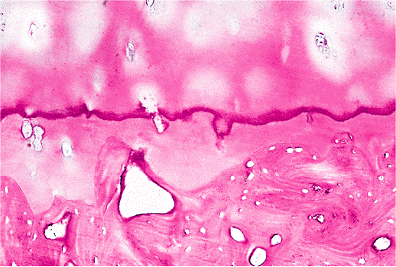 |
Figure 5.15 Photomicrograph of the deep and calcified layers of the articular cartilage. The deep layer is separated from the calcified layer by a basophilic line referred to as the tidemark, which represents the mineralizing front (H&E stain, 4 objective). |
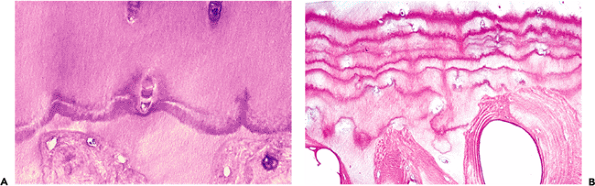 |
Figure 5.16 A. Photomicrograph demonstrating accelerated mineralization with a replicated tidemark. The mineralization front is almost certainly under cellular control; here, a chondrocyte is seen caught up in the tidemark (H&E stain, 25 objective). B. In most areas of normal cartilage, only one tidemark is observed. However, in an early stage of osteoarthritis, seen here, multiple tidemarks indicating rapid advance of the mineralization front can often be seen (H&E stain, 10 objective). |
At its base, adult articular cartilage is bordered by the subchondral bone plate, and the calcified cartilage tissue is keyed into the irregular surface of the underlying bone, somewhat like a jigsaw puzzle. Because the cartilage adjacent to the bone is calcified and has a rigidity similar to that of bone, the keying is rigid (Figure 5.17).
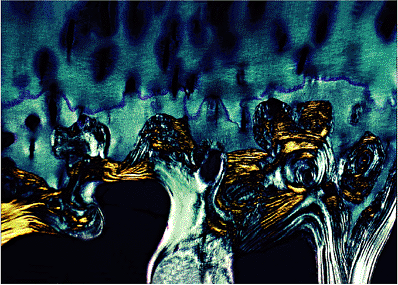 |
Figure 5.17 In this photomicrograph, taken with polarized light, the irregularity of the interface between the subchondral bone and the overlying calcified cartilage is obvious. The functional keying of the bone and cartilage depends on the two tissues having equal rigidity ( 4 objective). |
The insertions of ligaments and tendons are also calcified, and their insertions into the bone are effected by a similar keying. Because the insertions of ligament and tendons into the bone are generally studied in dry bone specimens, the bone markings we see are in fact the calcified portion of the ligament or tendon. Because the sites of such insertions are approximately the same from individual to individual, there is a tendency to think of them as static structures. However, since in the child growth is taking place continuously and in the adult bone turnover is continuously taking place (albeit slowly), it follows that the insertions of ligaments and tendons must participate in this dynamic process. (Our knowledge of anatomy is for the most part based upon the dissection of the dead; but, for morphology to be understood, time must be put into the equation. Life is characterized by continuous growth and change.)
The chondrocytes embedded in the cartilage matrix are responsible for synthesis and maintenance of the extracellular matrix of the tissue. The chondrocytes vary in size, shape, and number per unit volume of tissue, both from the superficial to the deep layers and in different anatomic locations (22) (Figure 5.18). Generally, cells at the cartilage surface are flatter, smaller, and orientated parallel with the cartilage surface. They also have a greater density than the cells deeper in the matrix (23). In the middle zones, chondrocytes are more spherical and arranged in columns. This vertical arrangement of cells probably reflects some interaction with the highly organized arrangement of collagen fibers in cartilage and suggests the possibility of movement of chondrocytes within the matrix substance as the collagen fibers are being laid down. (An analogy would be the precise organization of a spider's web that necessitates the movement of the spider.)
An interesting ultrastructural feature of chondrocytes is a nonmotile monocilium, which may have a mechanotransductory
P.107
function in regulation of matrix synthesis (24). This monocilium has been more frequently observed in young cartilage and reactive or reparative cartilage (25).
 |
Figure 5.18 A. Electron micrographs to illustrate the typical appearance of chondrocytes at the A. surface, B. mid-zone, and C. deep-zone of articular cartilage. At the surface the cells typically show more cell processes on the inferior surface. The Golgi and endoplasmic reticulum are less well developed than in the mid-zone. In the deep zone, the cells are degenerate, with disaggregated chromatin in the nucleus and vacuolization and fragmentation of the cytoplasm (all approximately 10,000). |
Chondrocytes are encased in a specialized layer of matrix distinctly different from the bulk of extracellular matrix. This layer is rich in proteoglycans, has some hyaluronic acid, and contains relatively little collagen. Around this paucicollagenous layer is a basketlike structure composed of cross-linked fibrillar collagen encapsulating the cell or sometimes groups of cells, and this provides a protective framework. Collagen type VI is found in this region.
In chondrocytes, mitochondria are sparse probably as a result of their comparatively low rates of oxygen consumption. Cells in the deeper uncalcified zone have the most prominent endoplasmic reticulum and Golgi apparatus, indicating active protein synthesis as well as sulfation of proteoglycan carbohydrate side chains. The cell membrane shows numerous short, as well as some longer, branched cytoplasmic processes, but they make no connection with the processes of other chondrocytes. In the extracellular matrix adjacent to the chondrocytes that lie above the tidemark, as in the hypertrophic zone of the growth plate, small
P.108
membrane-bound vesicles are visible. These may play a role in the calcification of cartilage matrix (26).
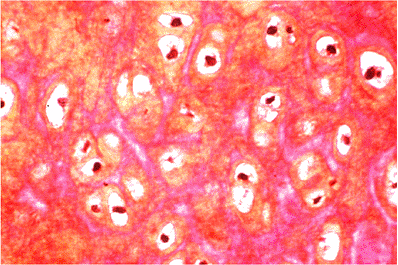 |
Figure 5.19 Photomicrograph of ear cartilage. Although the cells resemble those seen in hyalin cartilage, the matrix contains many elastic fibers that appear red in this section (phloxine and tartrazine stain, 25 objective). |
In addition to hyalin cartilage, of which articular cartilage is composed, two other forms of cartilage can be histologically recognized. Fibrocartilage is a tissue in which the matrix contains PG aggregates and a high proportion of type I collagen, the fibers of which are usually visible by transmitted light microscopy. Fibrocartilage may be found in the menisci of the knee, the annulus fibrosus, at the insertions of ligaments and tendons into the bone, and on the inner side of tendons as they angle around pulleys (e.g., at the malleoli). In all of these locations, the structures are subjected to compressive forces as well as tension. The second type of nonhyalin cartilage, elastic cartilage, contains a high proportion of elastin in the matrix and is found in the ligamentum flavum, external ear, and epiglottis (Figure 5.19). Compared to collagen, elastin has much greater elasticity; this is particularly important in the yellow ligaments, which make possible the flexion of the spinal canal.
Both the fibrocartilage and elastic cartilage incorporate the term cartilage because the cells are rounded and lie in lacunae, which give them a superficial microscopic resemblance to the cells of hyalin cartilage. However, the mechanical functions of these tissues are very different from those of hyalin cartilage. Hyalin cartilage is mainly subjected to and resists compressive forces, whereas both fibrocartilage and elastic cartilage function principally as resisters of tension, with some element of compression.
Cartilage Turnover and Articular Remodeling
Wolff's law states that both bone density and bone architecture correlate with the magnitude and direction of applied load. At the articular end of a bone, this implies that the subchondral bone trabeculae must also undergo a self-regulated modeling that maintains a joint shape capable of optimal load distribution. In other words, the shape of bones, including their articular ends, reflects a dynamic state that incorporates a feedback dependent on mechanical stress.
Endochondral ossification is an important mechanism for both growth and bone modeling. This is exemplified in the epiphyseal growth plate where calcified cartilage is invaded by blood vessels from the metaphyseal bone and is then replaced by bone tissue synthesized by osteoblasts lying close to the blood vessels.
Studies of adult joints have shown that replacement of the calcified layer of articular cartilage by bone tissue involves a similar process. Blood vessels from the subarticular bone penetrate the calcified cartilage, and new bone is laid down alongside the channels created by this process; thus the calcified cartilage is slowly replaced by new subarticular bone (Figure 5.20).
Replacement of the calcified layer of cartilage by bone might be expected to result in thinning and eventual disappearance of the calcified cartilage. However, histologic study of articular cartilage from subjects of various ages shows that the calcified cartilage remains much the same thickness throughout life. This is because the calcification front (tidemark) continues to advance into the noncalcified cartilage at a slow rate, which is in equilibrium with the rate of absorption of the calcified cartilage from the subarticular bone (27). Since the thickness of the articular cartilage does not significantly change during life, it can be postulated that articular cartilage is not a static tissue, as it was long believed to be. The extracellular matrix and the chondrocytes are being replaced throughout life; and, through these mechanisms, the joint undergoes continuous modeling. It seems likely that programmed cell death (apoptosis) plays an important role in this process in a similar way to what Mitrovic (28) has demonstrated in joint formation during limb development (29).
 |
Figure 5.20 In this photomicrograph, two vessels can be seen that have extended into the calcified layer of cartilage. Around the circumference of each of these vessels, a thin layer of lamellar bone can be appreciated. By means of continuing endochondral ossification, the articular bone end is continuously modeled (H&E stain, 10 objective). |
P.109
Heterogeneity of articular cartilage, including morphological, biochemical, and biomechanical variations, can be observed within different regions of a normal weight-bearing joint. Considerable variation in cartilage thickness over an articular surface is present in most joints. A variation in stiffness in different areas of the femoral head has been related both to PG content and to the amount of water held by the tissue (30). The stiffness of the cartilage is the main factor that determines stress in the tissue and, together with the thickness of the cartilage, has the largest effect on the stress in the calcified cartilage and underlying cancellous bone (31).
An example of the normal geographic variation in articular cartilage can be readily observed in the human tibial plateau, as well as in other animals, where there are distinct morphological differences between the articular cartilage that is covered by the meniscus and that which is not (32). These differences consist of a rough surface and soft matrix in the uncovered area as compared to the smooth, firm tissue that is covered by the meniscus. In adult human knee joints at autopsy, it has been found that articular cartilage that was not covered by meniscus (even as young as 17 years of age) always showed matrix softening and superficial fibrillation (33). The morphologic and biochemical findings in these two distinct articular areas as studied in the adult dog are summarized in Figure 5.21.
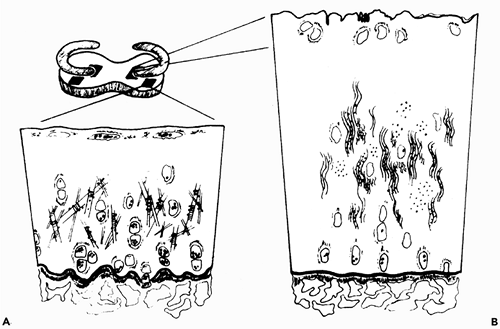 |
Figure 5.21 Morphological and chemical differences seen in the tibial articular cartilage under the meniscus, as compared with that not covered by the meniscus. A. In the covered area, the surface is smooth, and on the surface there is an amorphous electron-dense layer; the chondrocytes are flattened. With respect to lipid, there is an increased intracellular accumulation in all three layers. Increased accumulation of extracellular lipid is noted at the surface, and there are increased numbers of extracellular matrix vesicles in the deep zone. In electron microscopy sections, collagen appears as randomly oriented fibers of varying diameter but with thicker mean diameter than seen in B; there is regular binding of proteoglycan to the collagen fibrils; and the concentration of proteoglycans per wet weight is increased. The tidemark is irregular. B. In the uncovered area, the surface is irregular, there is a detached electron-dense layer, and the chondrocytes are rounded. The concentration of water per unit volume is increased. Collagen appears in wavy aggregated bundles with thinner mean diameters that vary but little from each other, and binding of proteoglycan to the collagen fibrils is ill defined. Proteoglycan can be extracted more easily from the cartilage matrix. The tidemark is smooth. In both the covered and the uncovered areas, the cell size is the same histologically, and there is the same amount of DNA per dry weight of cartilage tissue. (Modified from: Bullough PG, Yawitz PS, Tafra L, Boskey AL. Topographical variations in the morphology and biochemistry of adult canine tibial plateau articular cartilage. J Orthop Res 1985;3:1 16 with permission.) |
It has been postulated that these naturally occurring variations in matrix structure and mechanical properties are related to joint loading. In the normally functioning knee, load is transmitted through the meniscus and onto the tibial cartilage underlying the meniscus, whereas the exposed cartilage, that which is not covered by the meniscus, remains relatively unloaded. Similar areas of possible disuse atrophy have been described around the rim of the radial head, in
P.110
the roof of the acetabulum, and on the perifoveal and inferomedial aspects of the femoral head (34).
The extracellular matrix of the cartilage and of the other connective tissues is synthesized by their intrinsic cells under the control of both local and systemic factors. Both in vivo and in vitro studies have demonstrated that changes in the immediate environment of the joint lead to alterations of the cartilage matrix (35). Thus, immobilization or unloading of a joint results in decreased synthesis of glycosaminoglycans. Conversely, exercise appears to increase synthesis (36). These experimentally induced variations are in agreement with naturally observed topographic variations in joints that have been ascribed to normally occurring patterns of joint loading.
In general, it seems that low levels of mechanical stress (i.e., below the physiologic range) are associated with enhanced catabolic activity, whereas stress within the physiologic range is associated with more anabolic activity. Under conditions of supraphysiologic stress the chondrocytes are unable to adapt. In other words, there is a window of physiologic stress above or below which the chondrocytes cannot maintain an adequate functional matrix.
Although a number of factors have been implicated in the transduction of mechanical stimuli to metabolic events, the exact mechanism still remains unclear.
Histomorphogenesis of Articular Cartilage
Recent studies suggest that the histomorphogenesis of articular cartilage is regulated during skeletal development by the intermittent forces and motions acting at the site of diarthrodial joints. A key feature in this development is the formation of the superficial, transitional, radial, and zones through the cartilage thickness. The histomorphological and mechanical characteristics of these zones have been correlated with the distribution of pressures, deformations, and pressure-induced fluid flow created in vivo (31). However, unlike muscle and bone, the thickness of articular cartilage does not appear to adapt to mechanical stimulation (38).
Synovial Membrane
The synovial membrane lines the inner surface of the joint capsule and all other intra-articular structures, with the exception of articular cartilage and the meniscus. In addition to lining the joints, synovial membrane lines (a) the subcutaneous and subtendinous sacs known as bursae, which permit freedom of movement over a limited range for the structures adjacent to the bursae, and (b) the sheaths that form around tendons wherever they pass under ligamentous bands or through osseofibrous tunnels.
Synovial membrane consists of two components. The first of these is the cellular lining (or intimal layer) bounding the joint space. This surface is smooth, moist, and glistening, with a few small villi and fringelike folds. The second component is a subintimal, supportive, or backing layer (39).
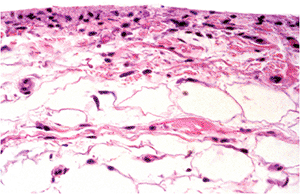 |
Figure 5.22 Photomicrograph of normal synovium. The ratio of fat-to-fibrous tissue varies depending on the joint and the location within the joint (H&E, 10 objective). |
Along the edge facing the synovial cavity, microscopic examination of synovial membrane reveals a single row or sometimes multiple rows of closely packed cells with large elliptical nuclei (Figure 5.22). Beneath the surface layer in the subintima, there is vascularized fibro-adipose tissue that contains some histiocytes, and mast cells.
Electron microscopic studies reveal two principal types of synovial lining cells, designated by Barland as Types A and B. (Many cells have features of both types and have been called intermediate.) The less common cell (Type A) has many of the features of a macrophage, and there is good evidence that it is structurally adapted for phagocytic functions. The more common Type B cells are richly endowed with rough endoplasmic reticulum, contain Golgi systems, and often show pinocytotic vesicles. Normal synovial intima contains 25% Type A and 75% Type B cells (40).
The synovial membrane has three principal functions: secretion of synovial fluid hyaluronate (Type B cells); phagocytosis of waste material derived from the various components of the joint (Type A cells); and regulation of the movement of solutes, electrolytes, and proteins from the capillaries into the synovial fluid, thus providing for the metabolic requirement of the joint chondrocytes and possibly also providing a regulatory mechanism for maintenance of the matrix through the role of various mediators.
Ligaments and Tendons
Ligaments, which are structures that join together two adjacent bones, are formed mainly of collagen. The arrangement of the collagen bundles within a particular ligament depend on the required movements within the joint; and,
P.111
because these movements are complex, the arrangement of the collagen bundles is similarly complex.
The collagen fibers of the ligament are calcified where they enter the bone, and the calcified portion of the ligament interdigitates and locks onto the underlying bone in the same way as does the calcified cartilage (vide supra). Just adjacent to the calcified portion of the ligament, the extracellular matrix of the ligament will be found to contain some proteoglycan; and, in association with this finding, the cells will be found to be contained within lacunaelike chondrocytes (41). The reason for this is that generally tendons enter the bone at an acute angle; for this reason a shear force acts on the ligament at its insertion and the normal tension in the ligament is complicated by an added compressive force (Figure 5.23).
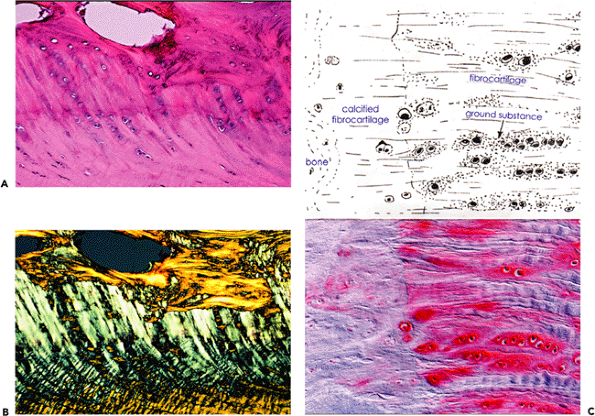 |
Figure 5.23 A. Photomicrograph of a ligamentous insertion using transmitted white light (H&E stain, 10 objective). B. Polarized light. The portion of the ligament that interfaces with the bone is calcified, and the edge of the calcified portion of the ligament is marked by a basophilic line (tidemark) that represents the mineralization front. Note the similarity with the bone cartilage interface illustrated in Figure 5.17. C. A higher powered view to demonstrate the rounded cells lying in lacunae, which are seen at the insertion site of both ligaments and tendons (fibrocartilaginous metaplasia). The red staining in the matrix indicates the presence of proteoglycans (safranin O stain, 25 objective). |
Tendons are specialized connective tissue structures that enable muscles to concentrate or extend their action. The Achilles tendon is a good example of a tendon that concentrates the power of several bulky muscles to one limited area of insertion; the long tendons of the hands and feet exemplify the function of extending the effect of distant muscles. Many muscles have no obvious tendinous insertions; for example the paravertebral and the gluteal muscles have short fan shaped fibrous insertions, which hardly justify their description as tendons.
The majority of cells within normal tendons are relatively inactive fibrocytes scattered in a sparse longitudinal pattern between the collagen bundles (Figure 5.24). There is a slight gradient in the cell population, the proximal (muscular) portion of the tendon being more cellular than its distal
P.112
insertion. The surfaces of the flexor tendons of the hand which glide within a synovial sheath, are covered by a single layer of synovial cells (the endotenon) and a similar layer covers the parietal surface of the fibrous tendon sheath (the epitenon). In the palm of the hand the tendons are covered by a fine vascular adventitia (paratenon) nourished by vessels from the deep palmar arch (42).
 |
Figure 5.24 Photomicrograph of the same field of a tendon that has been photographed in transmitted light in A and in polarized light in B. Both images demonstrate the scant and elongated fibroblasts lying between the dense parallel collagen bundles characteristic of tendon (H&E stain, 4 objective). |
The feeding arteries to the tendons of the hands and feet, both the vessels in the vinculae, and the vessels in the palmar or plantar paratenon are long, coiled vessels that can stretch as the tendons move. (The flexor tendons may glide some 6 cm between full flexion and full extension.)
Human tendons are poorly innervated. Apart from the nerve fibers associated with blood vessels and the free nerve endings in the region of the vinculae, there are very few nerve endings in the tendon proper.
Wherever a tendon turns a corner, or has to bend in association with neighboring joints, it is restrained under a pulley, or retinacular system, and is lubricated in this region by means of a synovial sheath. Because of the compression that occurs in these locations, just as at the insertion of the tendon or ligament, there is some cartilaginous metaplasia with the accumulation within the tissue of PGs that resist the compressive component of the load.
The Arthritic Joint
Clinical arthritis is the consequence of a breakdown in the joint's normal function; that is to say, it involves loss of capacity for the articulating surfaces to move over one another easily, loss of joint stability, and almost always pain.
The loss of freedom of motion may be the result of a change in joint shape that results in severe incongruities or, on the other hand, a change in the tissue matrices that affects their mechanical properties. Instability may result from alterations in ligamentous support and neuromuscular control. Pain may originate in the bone as a result of maldistribution of load; in the synovium as a result of reactive synovitis; or in the muscle as a consequence of reflex spasm.
Malfunction of a joint results from acute or chronic morbid conditions that produce either:
Anatomic alterations in the shape of the articulating surfaces (e.g., a transarticular or subarticular fracture, increased modeling activity, Paget's disease, or acromegaly) (43).
Loss of structural integrity of the cartilage tissue or of the support structures around the joint (e.g., by enzymatic destruction in inflammatory arthritis [septic or rheumatic arthritis] or, more commonly, traumatic injury).
Alterations in the mechanical properties of the tissue matrices making up the joint (e.g., brittle collagen as occurs in ochronosis).
During the past century, several types of arthritis have been well delineated on the basis of their characteristic clinical presentations and their morbid anatomy. These include the infectious arthritides, both granulomatous (TB) and pyogenic (septic); the metabolic arthritides (e.g., gout, pseudogout, and ochronosis); and the arthritis that complicates many cases of aseptic subchondral bone necrosis (44). The various rheumatic syndromes have been classified according to their clinical and immunologic characteristics; histologically, these inflammatory arthritides show chronic synovitis and a destructive pattern involving the bone, cartilage, and periarticular structures. However, they are difficult to differentiate from each other solely by microscopic examination.
Even when these various etiologies have been considered, there remain a large number of cases of arthritis affecting
P.113
especially certain small joints of the hands and feet and some larger joints, of which the hip and knee are most commonly involved. These cases, which run a chronic course, are essentially noninflammatory and usually occur in older individuals. The clinical presentation and morbid anatomy in these cases are similar enough for all of them to be classified under the general appellation of osteoarthritis or degenerative joint disease. In the majority of cases, the etiology is unclear; however, the important role of mechanical trauma is not in dispute. The onset of osteoarthritis in middle age can often be traced in sport-related injuries in adolescence and young adulthood. Repetitive impact loading, such as occurs in running on hard surfaces with poorly designed shoes, is recognized as contributing to knee arthritis. A study designed to understand the relationship between impactor energy and mass on injury modalities in the canine knee has shown that injuries were typically more frequent and more severe with the largest mass at each energy level. Histologic analysis of the patellae revealed cartilage injuries at low energy, with deep injuries in the underlying bone at higher energies (45).
Alteration in Shape
A change in joint shape is characteristic of most forms of arthritis. In the inflammatory arthritides tissue loss results from destruction. On the other hand, although bone and cartilage loss play an important part in the osteoarthritic process, it is the addition of new bone and cartilage in the form of osteophytes, particularly at the joint periphery and sometimes beneath the articular surface, that forms one of the characteristic features of the disease.
We now recognize that a change in joint shape either sudden, as with a fracture, or gradual, as in acromegaly or other metabolic disturbances such as Paget's disease may play an important role in the etiology of arthritis. In other words a change in the shape of the joint is an expected result of arthritis, but a change in shape may also be the cause of arthritis.
Tissue Response to Injury
Regardless of the etiology, joint injury is characterized by certain basic cellular and tissue responses.
There is usually macroscopic and microscopic evidence of both degeneration and of repair in the cells and in the extracellular matrix. (In the extracellular matrix, the changes may result from direct physical injury, from alteration in the cellular synthesis of the matrix, or from enzymatic breakdown of the matrix constituents. These changes are probably most apparent in the surface cartilage (46).
In the vascularized tissues, injury is followed by an acute and then by a chronic inflammatory response. As a result, the necrotic injured tissue is removed and replaced by proliferative vascular tissue (granulation tissue). The inflammatory response results in repair of injured tissue by fibrous scar. Independently of scarring, a second mode of repair involves regeneration of tissue similar to that which was injured originally. In nonvascularized tissue, such as cartilage, an inflammatory response and subsequent scarring cannot occur, but this does not preclude tissue regeneration. Note that cartilage injury always eventually invokes an inflammatory response since some vascularized tissues (i.e., bone and/or synovium) are inevitably involved.
Cartilage
Macroscopic (naked eye) evidence of injury to cartilage is evident only in the extracellular matrix, mainly the collagenous component; one of the earliest findings is a disruption of the collagen fibers at the surface, which, instead of being smooth, becomes rough and/or eroded (47). The local stresses and strains in the collagen fibrils, which cause the damage, cannot be determined dependably without taking the local arcadelike collagen-fibril structure into account (21). Three patterns of macroscopic alteration involving the cartilage surface and, to a variable degree, the underlying cartilage tissue can be identified: fibrillation (generally age related), erosion (ulceration), and cracking (probably trauma related) (48).
The term fibrillation is used to describe replacement of the normally smooth, shiny surface by a surface similar to cut velvet. This type of transformation can be observed both on very thick cartilage, such as the patella, and on very thin cartilage, such as that found in the interphalangeal joints. The pile of the fibrillated area may be short or shaggy. The junction between the fibrillated area and the adjacent normal appearing cartilage is morphologically usually well defined and generally distinct (Figure 5.25). A recent study has concluded that both microscopic magnetic resonance imaging and polarized light microscopy can detect quantitative changes in collagen fiber architecture in early osteoarthritis and also resolve topographical variation in cartilage microstructure of canine tibial plateau (49).
In this regard, continuing collaborative studies of morphology, including imaging studies, biochemistry, and biomechanics, are urgently needed.
For the morphologist, there appear to be two patterns of fibrillation. Well-defined areas of fibrillation affecting particular locations in certain joints are present in everyone from an early age (35). It is suggested by this author that these areas may be related to underloading of the cartilage. In osteoarthritic joints, there are areas of fibrillation that appear in different areas of the joint than those previously mentioned and that appear to be secondary to mechanical abrasion of the cartilage surface. The microscopic
P.114
characterization of these two distinct types of fibrillation is incomplete, but perhaps the latter is distinguished by deeper clefts and a greater tendency for the chondrocytes to form proliferating clones.
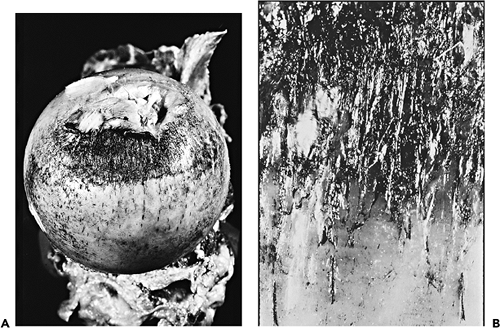 |
Figure 5.25 A. Photograph to demonstrate superficial fibrillation of the cartilage on the femoral head in the perifoveal region. The fibrillated cartilage has been highlighted by India ink. B. In a close up photograph, pin splits in the cartilage seem to follow the orientation of the collagen fibers in the fibrillated area. |
Cartilage ulceration, or solution of the surface, is characteristic of progressive degenerative changes in the joint. The base of the erosion appears initially to be either contoured or smooth. Tissue damage may eventually be so extensive as to completely denude the bone surface of its covering cartilage layer (eburnation).
The last form of structural lesion in this group, which is distinctly less common than either fibrillation or ulceration, is deep cracking of the cartilage. These cracks extend vertically deep into the cartilage and microscopically often have a deep horizontal component. Perhaps these result from severe impact loading (Figure 5.26).
In considering the pathogenesis of these three histologic types of cartilage matrix damage in the early stages of osteoarthritis, it is important to recognize that they may affect the opposed articular surfaces in different areas and to different degrees. This is in marked contrast to eburnation, in which both of the opposed surfaces are affected. It therefore appears that in many cases fibrillation and other cartilage alteration cannot be ascribed simply to abrasion.
An increase in the ratio of water to PG in the cartilage matrix leads to softening of the cartilage (chondromalacia), and this may be evidence of insufficient loading of the joint. Chondromalacia and fibrillation usually occur together, but chondromalacia may be present before there is any obvious gross evidence of fibrillation.
Injury at a cellular level is recognizable only microscopically. Necrosis can be identified when only the ghost outlines of the chondrocytes remain. This ghosting, usually scattered but focal in distribution, is a common finding in arthritis. Less often, all of the chondrocytes are seen to be necrotic (Figure 5.27).
Just as the effect of injury to the articular cartilage is reflected by the histologic response of both matrix and cells,
P.115
so too is the subsequent reparative cartilage regeneration. Within the preexisting cartilage matrix, there is focal cell proliferation with clumps, or clones, of chondrocytes. When the tissue is stained with toluidine blue, there is often intense metachromasia of the matrix around these clumps of proliferating chondrocytes, evidence of increased PG synthesis. This process can be thought of as intrinsic repair (50) (Figure 5.28).
 |
Figure 5.26 Photomicrograph demonstrating deep cracking of the cartilage matrix. The lesion shown is characteristic of a blisterlike lesion, which is seen in many cases of chondromalacia patellae (H&E stain, 4 objective). |
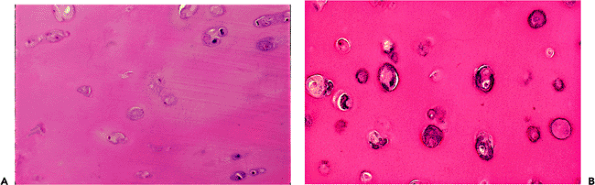 |
Figure 5.27 A. A photomicrograph to demonstrate focal chondrocyte necrosis. In cases of degenerative arthritis, focal areas of necrosis (such as seen here) are common. Rarely, the necrosis is extensive. In inflammatory arthritis, chondrocyte necrosis is also common and often associated with an irregular lysis of the matrix around the necrotic cells, the so-called Weichselbaum's lacunae (H&E stain, 10 objective). B. Photomicrograph to demonstrate focal calcification around necrotic chondrocytes in the deep zone of the cartilage (H&E stain, 25 objective). |
In a damaged joint, cartilage repair may also be initiated from either or both of two possible sites, either the joint margin or the subchondral bone. Extrinsic repair of cartilage, which develops from the joint margin, can be seen as a cellular layer of cartilage extending over, and sometimes dissecting into, the existing cartilage. This extrinsically repaired cartilage is usually much more cellular than the preexisting articular cartilage, and the chondrocytes are evenly distributed throughout the matrix (Figure 5.29).
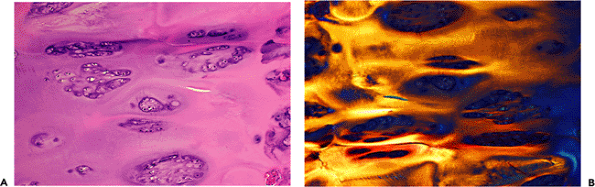 |
Figure 5.28 A. Photomicrograph to demonstrate clones of regenerating chondrocytes. Note the basophilia around the clones, which correspond to increased proteoglycan synthesis by the cells (H&E stain, 10 objective). B. When examined by polarized light, the proliferating clones are visibly displacing the existing collagen matrix. |
On microscopic examination this type of repair cartilage can easily be overlooked. However, examination under polarized light will clearly demonstrate the discontinuity between the collagen network of the repair cartilage and that of the preexisting cartilage (Figure 5.30), as well as the denser thicker collagen fibers of the repair cartilage.
In arthritic joints in which loss of the articular cartilage has denuded the underlying bone, especially in cases of osteoarthritis, there are frequently small pits in the bone surface from which protrude small nodules of firm white
P.116
P.117
tissue. On microscopic examination, these nodules have the appearance of fibrocartilage arising in the marrow spaces of the subchondral bone. They may extend over the previously denuded surface to form a more or less continuous layer of repair tissue. Most specimens obtained from cases of osteoarthritis reveal both intrinsic and extrinsic repair of cartilage (51).
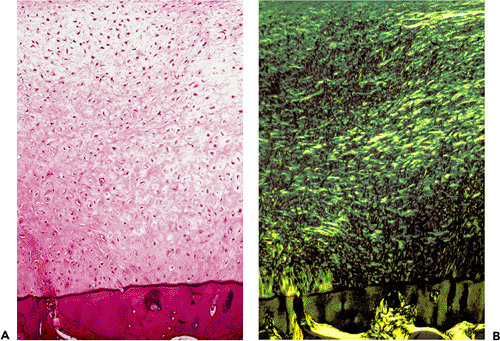 |
Figure 5.29 A. A section through the articular surface of an arthritic joint demonstrates extrinsic reparative fibrocartilage, which extends to the tidemark of the original articular hyaline cartilage (H&E stain, 10 objective). B. The same field photographed with polarized light shows the discontinuity of the collagen between the calcified zone and the reparative cartilage. |
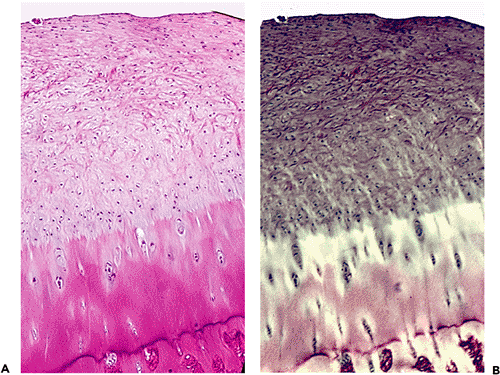 |
Figure 5.30 A. Photomicrograph showing reparative cartilage extending over preexisting damaged cartilage (H&E stain, 4 objective). B. Same field photographed with polarized light. |
Bone
Arthritis is a disease that affects not only the articular cartilage, but also the underlying bone and the structures around the joint.
As the articular cartilage is eroded from the surface, the underlying bone is subjected to increasingly localized overloading. In subarticular bone that has been denuded, there is proliferation of osteoblasts and formation of new bone, which occurs both on the surfaces of existing intact trabeculae and around microfractures (52) (Figure 5.31). In x-rays of arthritic joints, this new bone appears as increased density or sclerosis.
A further result of increased local stress is that the bone at the articulating surface is likely to undergo focal pressure necrosis (Figure 5.32). (This superficial necrosis is different both in its etiology and pathogenesis from that associated with primary subchondral infarction, which itself leads to secondary osteoarthritis. However, in clinical practice, differentiation between the two may be difficult, especially in the late stages of primary subchondral infarction.) (53)
Subarticular cysts are usually seen only where the overlying cartilage is absent. Such cysts are common in cases of osteoarthritis and are believed to result from transmission of intra-articular pressure through defects in the articulating bony surface into the marrow spaces of the subchondral bone (54). The cysts increase in size until the pressure within them is equal to the intra-articular pressure.
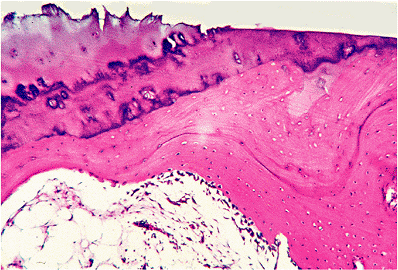 |
Figure 5.31 Photomicrograph of the edge of an eburnated area of bone in a case of osteoarthritis. There is a very prominent layer of osteoblasts covering the sclerotic bone that underlies the area denuded of cartilage (H&E stain, 4 objective). |
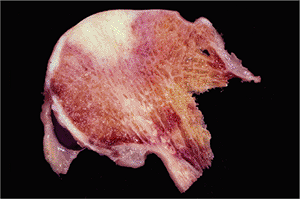 |
Figure 5.32 A section through an osteoarthritic femoral head shows a large wedge shaped area of necrosis of the superior portion of the head. |
Cysts may also occur because of focal tissue necrosis (55). (In cases of arthritis due to rheumatoid disease or gout, periarticular radiologic cysts may be associated with erosion of the marginal subchondral bone by the diseased synovium.)
Separated fragments of bone and cartilage from a damaged joint surface may become incorporated into the synovial membrane and digested, or they may remain free as loose bodies in the joint cavity. Under certain circumstances, proliferation of cartilage cells occurs on the surface of these loose bodies and consequently they grow larger (Figure 5.33). As they grow, their centers become necrotic and calcified. In histologic sections it is possible to visualize periodic extension of this central calcification in the form of concentric rings, which increase in number as the loose body grows larger. Sometimes the loose bodies reattach to the synovial membrane at a later stage, in which case they are invaded by blood vessels. Endochondral ossification then occurs, and the loose bodies again develop a viable bony core.
There is some degree of loose body formation in many cases of arthritis, but they are especially prominent in Charcot's joints and in other types of rapidly destructive osteoarthritis. Occasionally, in cases of osteoarthritis, the loose bodies are so numerous that they must be distinguished from those that occur in primary synovial chondromatosis (56).
Ligaments
Microscopic evidence both of lacerations and of repair by scar tissue is common in the ligamentous and capsular tissue around an arthritic joint. These changes are readily recognized by the use of polarized microscopy, where the
P.118
alterations in the organization of the collagen are made very clear (Figure 5.34). Whether these lacerations preceded the arthritic process or whether they are a consequence of it cannot usually be determined by microscopic examination.
 |
Figure 5.33 A. Gross photograph of multiple loose bodies in a case of osteoarthritis of the hip joint. B. Low-power photograph of a cross section of a loose body showing concentric growth rings (H&E stain, 1 objective). C. Photomicrograph showing crowded proliferating chondrocytes and a growth ring (H&E stain, 4 objective). D. Photomicrograph to show benign proliferating chondrocytes (H&E stain, 25 objective). |
Synovial Membrane
Injury and breakdown of cartilage and bone result in increased amounts of breakdown product and particulate debris within the joint cavity. This is removed from the synovial fluid by phagocytic cells (the Type A cells) of the synovial membrane. In consequence, the membrane becomes both hypertrophic and hyperplastic, and the breakdown products of the cartilage and bone matrix frequently evoke an inflammatory response (Figure 5.35).
For this reason, some degree of chronic inflammation can be expected in the synovial membrane of arthritic joints, even when the injury has been purely a mechanical one. Inflammation is especially prominent where there has been rapid breakdown of the articular components as evidenced by the presence in the synovium of bone and cartilage detritus.
Histologic studies have shown that there may be a similarity between the degree of inflammatory response as seen in some cases of severe osteoarthritis and that of rheumatoid arthritis (57). However, in osteoarthritis the synovial inflammation is likely to be the result of cartilage breakdown, whereas in rheumatoid arthritis the synovial inflammation is the cause of cartilage breakdown.
Extension of the hyperplastic synovium onto the articular surface of the joint (i.e., a pannus) is a common finding even in osteoarthritis, particularly in the hip (Figure 5.36). However, the extent and the aggressiveness of this pannus with respect to underlying cartilage destruction is much less marked in osteoarthritis than in rheumatoid arthritis.
Since, under normal conditions, the synovial membrane is responsible for the nutrition of articular cartilage, it might be expected that the chronically inflamed and scarred synovial membrane of an arthritic joint would function less effectively than that of a normal joint. Disturbance in synovial nutrient function, as well as increased enzymatic
P.119
activity, may very well contribute to the chronicity of the arthritic process.
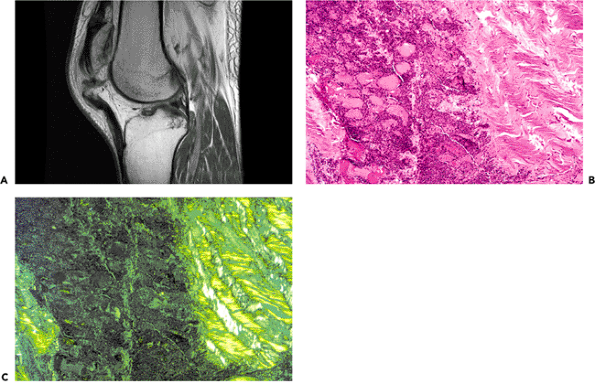 |
Figure 5.34 A. A magnetic resonance image of a knee shows rupture of the patellar ligament. B. Photomicrograph to demonstrate an area in a ligament where a laceration has occurred. The well-oriented collagen of the lacerated ligament is clearly demarcated from the resultant defect, which can be seen to have been filled with a vascularized cellular fibrous scar tissue (H&E stain, 10 objective). C. Same field photographed with polarized light. |
The hypertrophied and hyperplastic synovium associated with arthritis is also likely to be traumatized as it extends into the joint cavity. Evidence of bleeding into the joint, with subsequent hemosiderin staining of the synovial membrane, is a common histologic finding and may occasionally be marked. When this is the case, and despite their similar color, the orange-brown staining of the fine villous synovium seen at operation should not be confused with the swollen papillary synovium of pigmented villonodular synovitis.
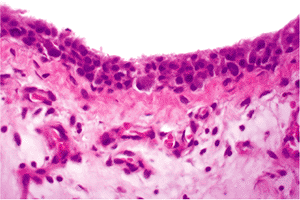 |
Figure 5.35 Photomicrograph of the synovium removed from the joint of a patient with a moderate degree of osteoarthritis reveals not only a hypertrophy of the synovial lining cells, but also hyperplasia that has resulted in a piling up of the synoviocytes. In the subsynovial tissue, there is increased vascularity and a mild chronic inflammatory infiltrate (H&E stain, 25 objective). |
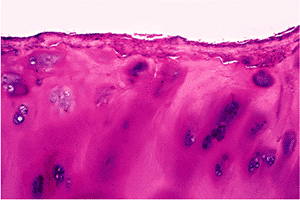 |
Figure 5.36 Photomicrograph of a portion of the articular surface of a femoral head in a case of osteoarthritis. A fibrous pannus extends over the articular surface (H&E stain, 10 objective). |
P.120
Synovial Fluid
Examination of synovial fluid is extremely helpful in the diagnosis of arthritis, both for determining the cause and the stage of the disease. Whatever the cause of arthritis, the synovial fluid is altered.
Normal synovial fluid, a dialysate of plasma to which hyaluronic acid produced by the Type B cells of the synovial lining is added, is viscous, pale yellow, and clear. Even in large joints the volume is small.
In cases of inflammatory arthritis, there is an increased volume of synovial fluid with a high count of inflammatory cells. The amount of hyaluronic acid is markedly diminished, leading to a typical decrease in viscosity. On the other hand, in degenerate forms of arthritis the amount of hyaluronic acid is increased, resulting in an extremely viscous fluid. There is also an increase in volume, although not to the same degree as that which is seen in the inflammatory arthritides.
References
1. Hammond BT, Charnley J. The sphericity of the femoral head. Med Biol Eng 1967;5:445 453.
2. Bullough P, Goodfellow J, O'Conner J. The relationship between degenerative changes and load-bearing in the human hip. J Bone Joint Surg Br 1973;55:746 758.
3. Bullough PG, Walker PS. The distribution of load through the knee joint and its possible significance to the observed patterns of articular cartilage breakdown. Bull Hosp Joint Dis 1976;37:110 123.
4. Hunter W. On the structure and disease of articulating cartilages. Phil Trans 1743;42:514 521.
5. Eyre DR. Collagens and cartilage matrix homeostasis. Clin Orthop Relat Res 2004; 427(suppl):S118 S122.
6. Mayne R, Irwin MH. Collagen types in cartilage. In: Kuettner KE, Schleyerbach R, Hascall VC, eds. Articular Cartilage Biochemistry. New York: Raven Press; 1986:23.
7. Eyre DR, Wu JJ, Woods P. Cartilage-specific collagens: structural studies. In: Kuettner KE, Schleyerbach R, Peyron JG, Haskell VC, eds. Articular Cartilage and Osteoarthritis. New York: Raven Press; 1992:119 132.
8. Watanabe H, Yamada Y, Kimata K. Roles of aggrecan, a large chondroitin sulfate proteoglycan, in cartilage structure and function. J Biochem (Tokyo) 1998;124:687 693.
9. Kiani C, Chen L, Wu YJ, Yee AJ, Yang BB. Structure and function of aggrecan. Cell Res 2002;12:19 32.
10. Roughley PJ. Articular cartilage and changes in arthritis: noncollagenous proteins and proteoglycans in the extracellular matrix of cartilage. Arthritis Res 2001;3:342 347.
11. Smith MM, Gosh P. Experimental models of osteoarthritis. In: Moskowitz RW, Howell DS, Goldberg VM, Mankin HJ, eds. Osteoarthritis:Diagnosis and Medical/Surgical Management. 3rd ed. Philadelphia: WB Saunders; 2001:171 200.
12. Kempson GE. Mechanical properties of articular cartilage. In: Freeman MAR, ed. Adult Articular Cartilage. 2nd ed. London: Pitman Medical; 1973.
13. Van der Korst JK, Skoloff L., Miller EJ. Senescent pigmentation of cartilage and degenerative joint disease. Arch Pathol 1968;86:40 47.
14. Maroudas A, Bullough P, Swanson SA, Freeman MAR. The permeability of articular cartilage. J Bone Joint Surg Br 1968;50:166 177.
15. Hultkranz W. Ueber die Spaltrichtungen der Gelenkknorpel. Verhandlungen der Anatomischen Gesellschaft. 1898;12:248.
16. Bullough P, Goodfellow J. The significance of the fine structure of articular cartilage. J Bone Joint Surg Br 1968;50:852 857.
17. Benninghoff A. Form und Bau der Gelenkknorpel in ihren Beziehungen zur Funktion. II. Der Aufbau des Gelenkknorpels in seinen Beziehungen zur Funktion. Z Zellforsch Mikrosk Anat 1925;2:783 862.
18. Muir H, Bullough P, Maroudas A. The distribution of collagen in human articular cartilage with some of its physiological implications. J Bone Joint Surg Br 1970;52:554 563.
19. Wilson W, van Donkelaar CC, van Rietbergen B, Ito K, Huiskes R. Stresses in the local collagen network of articular cartilage: a poroviscoelastic fibril-reinforced finite element study. J Biomech 2004;37:357 366.
20. Bullough PG, Munuera L, Murphy J, Weinstein AM. The strength of the menisci of the knee as it relates to their fine structure. J Bone Joint Surg Br 1970;52:564 567.
21. Maroudas A, Evans H, Almeida L. Cartilage of the hip joint. Topographical variation of glycosaminoglycan content in normal and fibrillated tissue. Ann Rheum Dis 1973;32:1 9.
22. Stockwell RA, Meachim G. The chondrocytes. In: Freeman MAR, ed. Adult Articular Cartilage. London: Pitman Medical; 1973.
23. Stockwell RA. The interrelationship of cell density and cartilage thickness in mammalian articular cartilage. J Anat 1971;109:411 421.
24. Wilsman NJ. Cilia of adult canine articular chondrocytes. J Ultrastruct Res 1978;64:270 281.
25. Stockwell RA. The ultrastructure of cartilage canals and the surrounding cartilage in the sheep fetus. J Anat 1971;109(pt 3):397 410.
26. Anderson HC. Calcification processes. Pathol Annu 1980;15(pt 2):45 75.
27. Boskey AL, Bullough PG, Dmitrovsky E. The biochemistry of the mineralization front. Metab Bone Dis Relat Res 1980;2S:61 67.
28. Mitrovic D. Regression of normally constituted articular cavities in paralyzed chick embryos [in French]. C R Acad Sci Hebd Seances Acad Sci D 1972;274:288 291.
29. Mori C, Nakamura N, Kimura S, Irie H, Takigawa T, Shiota K. Programmed cell death in the interdigital tissue of the fetal mouse limb is apoptosis with DNA fragmentation. Anat Rec 1995;242:103 110.
30. Kempson GE. Mechanical Properties of Human Articular Cartilage. [doctoral thesis]. London:University of London; 1970.
31. Dar FH, Aspden RM. A finite element model of an idealized diarthrodial joint to investigate the effects of variation in the mechanical properties of the tissues. Proc Inst Mech Eng H 2003;217:341 348.
32. Bullough PG, Yawitz PS, Tafra L, Boskey AL. Topographical variations in the morphology and biochemistry of adult canine tibial plateau articular cartilage. J Orthop Res 1985;3:1 16.
33. Bennet GA, Waine H, Bauer W. Changes in the Knee Joint at Various Ages: With Particular Reference to the Nature and Development of Degenerative Joint Disease. New York: Commonwealth Fund; 1942.
34. Goodfellow JW, Bullough PG. The pattern of ageing of the articular cartilage of the elbow joint. J Bone Joint Surg Br 1967;49:175 181.
35. Palmoski MJ, Brandt KD. Effects of static and cyclic compressive loading on articular cartilage plugs in vitro. Arthritis Rheum 1984;27:675 681.
36. Palmoski M, Perricone E, Brandt KD. Development and reversal of a proteoglycan aggregation defect in normal canine knee cartilage after immobilization. Arthritis Rheum 1979;22:508 517.
37. Wong M, Carter DR. Articular cartilage functional histomorphology and mechanobiology: a research perspective. Bone 2003;33:1 13.
38. Eckstein F, Faber S, Muhlbauer R, et al. Functional adaptation of human joints to mechanical stimuli. Osteoarthritis Cartilage 2002;10:44 50.
39. Henderson B, Pettipher ER. The synovial lining cell: biology and pathobiology. Semin Arthritis Rheum 1985;15:1 32.
40. Barland P, Novikoff AB, Hamerman D. Electron microscopy of the human synovial membrane. J Cell Biol 1962;14:207 220.
41. Cooper RR, Misol S. Tendon and ligament insertion. A light and electron microscopic study. J Bone Joint Surg Am 1970;52:1 20.
P.121
42. Lundborg G, Myrhage R. The vascularization and structure of the human digital tendon sheath as related to flexor tendon function. An angiographic and histological study. Scand J Plast Reconstr Surg 1977;11:195 203.
43. Johanson NA. Endocrine arthropathies. Clin Rheum Dis 1985;11:297 323.
44. Bullough PG, DiCarlo EF. Subchondral avascular necrosis: a common cause of arthritis. Ann Rheum Dis 1990;49:412 420.
45. Atkinson PJ, Ewers BJ, Haut RC. Blunt injuries to the patellofemoral joint resulting from transarticular loading are influenced by impactor energy and mass. J Biomech Eng 2001;123:293 295.
46. Rieppo J, Toyras J, Nieminen MT, et al. Structure-function relationships in enzymatically modified articular cartilage. Cells Tissues Organs 2003;175:121 132.
47. Collins DH. The Pathology of Articular and Spinal Diseases. London: Edward Arnold, 1949.
48. Heine J. ber die Arthritis deformans. Virchows Arch Path Anat 1926;260:521 663.
49. Alhadlaq HA, Xia Y, Moody JB, Matyas JR. Detecting structural changes in early experimental osteoarthritis of tibial cartilage by microscopic magnetic resonance imaging and polarised light microscopy. Ann Rheum Dis 2004;63:709 717.
50. Nakata K, Bullough PG. The injury and repair of human articular cartilage: a morphological study of 192 cases of coxarthrosis. Nippon Seikeigeka Gakkai Zasshi 1986;60:763 775.
51. Macys JR, Bullough PG, Wilson PD Jr. Coxarthrosis: a study of the natural history based on a correlation of clinical, radiographic, and pathologic findings. Semin Arthritis Rheum 1980;10:66 80.
52. Christensen SB. Osteoarthrosis. Changes of bone, cartilage and synovial membrane in relation to bone scintigraphy. Acta Orthop Scand Suppl 1985;214:1 43.
53. Franchi A, Bullough PG. Secondary avascular necrosis in coxarthrosis: a morphologic study. J Rheumatol 1992;19:1263 1268.
54. Landells JW. The bone cysts of osteoarthritis. J Bone Joint Surg Br 1953;35 B:643 649.
55. Rhaney K, Lamb DW. The cysts of osteoarthritis of the hip: a radiological and pathological study. J Bone Joint Surg Br 1955;37-B:663 675.
56. Villacin AB, Brigham LN, Bullough PG. Primary and secondary synovial chondrometaplasia: histopathologic and clinicoradiologic differences. Hum Pathol 1979;10:439 451.
57. Ito S, Bullough PG. Synovial and osseous inflammation in degenerative joint disease and rheumatoid arthritis of the hip. Histometric study. Proceedings of the 25th Annual ORS 1979;199.