10. The 3 ends of mRNAs are generated by cleavage
22.9 The 3′ ends of polI and polIII transcripts are generated by termination |
 |
Figure 22.28 When a 3 F end is generated by termination, RNA polymerase and RNA are released at a discrete (terminator) sequence in DNA. |
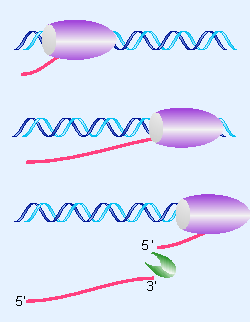 |
Figure 22.29 When a 3 F end is generated by cleavage, RNA polymerase continues transcription while an endonuclease cleaves at a defined sequence in the RNA. |
3′ends of RNAs can be generated in two ways. Some RNA polymerases terminate transcription at a defined (terminator) sequence in DNA, as shown in Figure 22.28. Other RNA polymerases do not show discrete termination, but continue past the site corresponding to the 3′end, which is generated by cleavage of the RNA by an endonuclease, as shown in Figure 22.29.
Information about the termination reaction for eukaryotic RNA polymerases is much less detailed than our knowledge of initiation. RNA polymerases I and III have discrete termination events (like bacterial RNA polymerase), but it is not clear whether RNA polymerase II usually terminates in this way.
For RNA polymerase I, the sole product of transcription is a large precursor that contains the sequences of the major rRNA. The precursor is subjected to extensive processing. Termination occurs at a discrete site >1000 bp downstream of the mature 3′ end, which is generated by cleavage. Termination involves recognition of an 18 base terminator sequence by an ancillary factor.
With RNA polymerase III, transcription in vitro generates molecules with the same 5′ and 3′ ends as those synthesized in vivo. The termination reaction resembles intrinsic termination by bacterial RNA polymerase. Termination usually occurs at the second U within a run of 4 U bases, but there is heterogeneity, with some molecules ending in 3 or even 4 U bases. The same heterogeneity is seen in molecules synthesized in vivo, so it seems to be a bona fide feature of the termination reaction.
Just like the prokaryotic terminators, the U run is embedded in a G PC-rich region. Although sequences of dyad symmetry are present, they are not needed for termination, since mutations that abolish the symmetry do not prevent the normal completion of RNA synthesis. Nor are any sequences beyond the U run necessary, since all distal sequences can be replaced without any effect on termination.
The U run itself is not sufficient for termination, because regions of 4 successive U residues exist within transcription units read by RNA polymerase III. (However, there are no internal U5 runs, which fits with the greater efficiency of termination when the terminator is a U5 rather than U4 sequence.) The critical feature in termination must therefore be the recognition of a U4 sequence in a context that is rich in G PC base pairs.
How does the termination reaction occur? It cannot rely on the weakness of the rU-dA RNA-DNA hybrid region that lies at the end of the transcript, because often only the first two U residues are transcribed. Perhaps the G PC-rich region plays a role in slowing down the enzyme, but there does not seem to be a counterpart to the hairpin involved in prokaryotic termination. We remain puzzled how the enzyme can respond so specifically to such a short signal. And in contrast with the initiation reaction, which RNA polymerase III cannot accomplish alone, termination seems to be a function of the enzyme itself.