5. RNA polymerase III uses both downstream and upstream promoters
20.5 RNA polymerase III uses both downstream and upstream promoters |
| Key terms defined in this section |
| Preinitiation complex in eukaryotic transcription describes the assembly of transcription factors at the promoter before RNA polymerase binds. |
Recognition of promoters by RNA polymerase III illustrates strikingly the relative roles of transcription factors and the polymerase enzyme. The promoters fall into two general classes that are recognized in different ways by different groups of factors. The promoters for 5S and tRNA genes are internal; they lie downstream of the startpoint. The promoters for snRNA (small nuclear RNA) genes lie upstream of the startpoint in the more conventional manner of other promoters. In both cases, the individual elements that are necessary for promoter function consist exclusively of sequences recognized by transcription factors, which in turn direct the binding of RNA polymerase.
Before the promoter of 5S RNA genes was identified in X. laevis, all attempts to identify promoter sequences assumed that they would lie upstream of the startpoint. But deletion analysis showed that the 5S RNA product continues to be synthesized when the entire sequence upstream of the gene is removed!
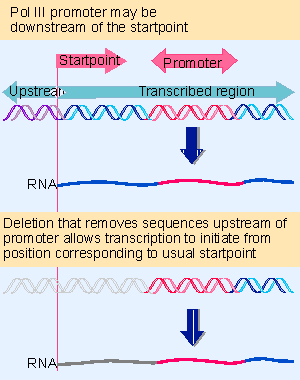 |
Figure 20.5 Deletion analysis shows that the promoter for 5S RNA genes is internal; initiation occurs a fixed distance (~55 bp) upstream of the promoter. |
When the deletions continue into the gene, a product very similar in size to the usual 5S RNA continues to be synthesized so long as the deletion ends before base +55. Figure 20.5 shows that the first part of the RNA product corresponds to plasmid DNA; the second part represents the segment remaining of the usual 5S RNA sequence. But when the deletion extends past +55, transcription does not occur. So the promoter lies downstream of position +55, but causes RNA polymerase III to initiate transcription a more or less fixed distance away.
When deletions extend into the gene from its distal end, transcription is unaffected so long as the first 80 bp remain intact. Once the deletion cuts into this region, transcription ceases. This places the downstream boundary position of the promoter at about position +80.
So the promoter for 5S RNA transcription lies between positions +55 and +80 within the gene. A fragment containing this region can sponsor initiation of any DNA in which it is placed, from a startpoint ~55 bp farther upstream. (The wild-type startpoint is unique; in deletions that lack it, transcription initiates at the purine base nearest to the position 55 bp upstream of the promoter. (641, 642))
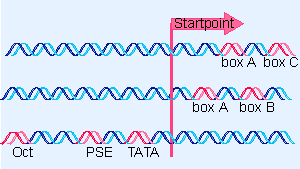 |
Figure 20.6 Promoters for RNA polymerase III may consist of bipartite sequences downstream of the startpoint, with boxA separated from either boxC or boxB. Or they may consist of separated sequences upstream of the startpoint (Oct, PSE, TATA). |
The structures of three types of promoters for RNA polymerase III are summarized in Figure 20.6. There are two types of internal promoter. Each contains a bipartite structure, in which two short sequence elements are separated by a variable sequence. Type 1 consists of a boxA sequence separated from a boxC sequence, and type 2 consists of a boxA sequence separated from a boxB sequence. The distance between boxA and boxB in a type 2 promoter can vary quite extensively, but the boxes usually cannot be brought too close together without abolishing function. We discuss the organization of the upstream type of promoter later.
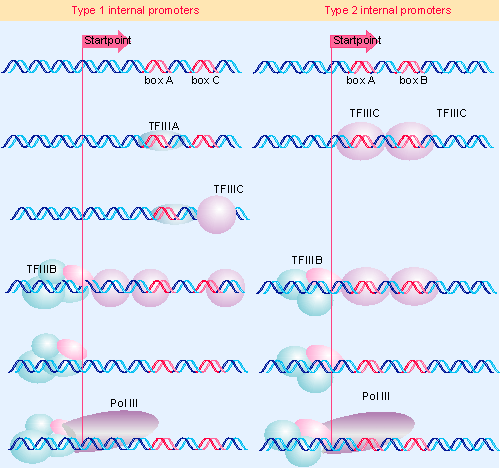 |
Figure 20.7 Initiation via the internal pol III promoters involves the assembly factors TFIIIA and TFIIIC, the initiation factor TFIIIB, and RNA polymerase III. |
Figure 20.7 summarizes the stages of reaction at internal promoters. Three accessory factors are involved. TFIIIA is a member of an interesting class of zinc finger proteins that we discuss later. TFIIIB consists of three subunits (see below). TFIIIC is a large protein complex (>500 kD), comparable in size to RNA polymerase itself, and containing at least 5 subunits.
We do not fully understand all the interactions that occur at the pol III promoters, but the principle is clear. At type 2 promoters (on the right side of the figure), TFIIIC recognizes boxB, but binds to a more extensive region including both boxes A and B. At type 1 promoters (on the left side of the figure), TFIIIA binds to a sequence that includes boxC, and this is required to enable TFIIIC to bind. In both cases, the binding of TFIIIC in turn enables TFIIIB to bind to a sequence surrounding the startpoint.
A crucial feature in defining the roles of the factors is that, at this point, TFIIIA and TFIIIC can be removed from the promoter (by high salt concentration in vitro) without affecting the initiation reaction. TFIIIB remains bound in the vicinity of the startpoint and its presence is sufficient to allow RNA polymerase III to bind at the startpoint. So TFIIIB is the only true initiation factor required by RNA polymerase III. TFIIIA and TFIIIC are assembly factors, whose role is to assist the binding of TFIIIB at the right location. This sequence of events explains how the promoter boxes downstream can cause RNA polymerase to bind at the startpoint, farther upstream.
So TFIIIB functions as a "positioning factor," responsible for localizing RNA polymerase correctly. Like SL1 at the pol I promoter, it resembles a sigma factor, in lacking the ability to bind DNA by itself, but being able to bind in conjunction with other proteins. TFIIIB includes the same protein, TBP, that is present in SL1, and also in the corresponding transcription factor (TFIID) for RNA polymerase II (643). It also contains Brf, which is related to the factor TFIIB that is used by RNA polymerase II). The third subunit is called B"; it is dispensable if the DNA duplex is partially melted, which suggests that its function is to initiate the transcription bubble (945). The role of B" may be comparable to the role played by sigma factor in bacterial RNA polymerase (see 9.7 Substitution of sigma factors may control initiation).
Although the ability to transcribe these genes is conferred by the internal promoter, changes in the region immediately upstream of the startpoint can alter the efficiency of transcription.
The upstream region has a more important role in the third class of polymerase III promoters. In the example shown in Figure 20.6, there are three upstream elements. These elements are also found in promoters for snRNA genes that are transcribed by RNA polymerase II. (Genes for some snRNAs are transcribed by RNA polymerase II, while others are transcribed by RNA polymerase III.) The upstream elements function in a similar manner in promoters for both polymerases II and III.
Initiation at an upstream promoter for RNA polymerase III can occur on a short region that immediately precedes the startpoint and contains only the TATA element. However, efficiency of transcription is much increased by the presence of the PSE and OCT elements. The factors that bind at these elements interact cooperatively. (The PSE element may be essential at promoters used by RNA polymerase II, whereas it is stimulatory in promoters used by RNA polymerase III; its name stands for proximal sequence element.)
The TATA element appears to confer specificity for the type of polymerase (II or III) that is recognized by an snRNA promoter. It is recognized by a factor that includes the TBP, which actually recognizes the sequence in DNA. The TBP is associated with other proteins, which are specific for the type of promoter. The function of TBP and its associated proteins is to position the RNA polymerase correctly at the startpoint. We discuss this in more detail for RNA polymerase II in the next section.
The factors work in the same way for both types of promoters for RNA polymerase III. The factors bind at the promoter before RNA polymerase itself can bind. They form a preinitiation complex that directs binding of the RNA polymerase. RNA polymerase III does not itself recognizes the promoter sequence, but binds adjacent to factors that are themselves bound just upstream of the startpoint. For the type 1 and type 2 internal promoters, the assembly factors ensure that TFIIIB (which includes TBP) is bound just upstream of the startpoint, to provide the positioning information. For the upstream promoters, transcription factors that directly recognize the upstream sites form a complex (including TBP) that is recognized by RNA polymerase III. So irrespective of the location of the promoter sequences, factor(s) are bound close to the startpoint in order to direct binding of RNA polymerase III (for review see 220).
This section updated 2-3-2000
| Reviews | |
| 220: | Geiduschek, E. P. and Tocchini-Valentini, G. P. (1988). Transcription by RNA polymerase III. Ann. Rev. Biochem 57, 873-914. |
| Research | |
| 641: | Sakonju, S., Bogenhagen, D. F., and Brown, D. D. (1980). A control region in the center of the 5S RNA gene directs specific initiation of transcription: I the 5?/FONT> border of the region. Cell 19, 13-25. |
| 642: | Bogenhagen, D. F., Sakonju, S., and Brown, D. D. (1980). A control region in the center of the 5S RNA gene directs specific initiation of transcription: II the 3?/FONT> border of the region. Cell 19, 27-35. |
| 643: | Kassavatis, G. A., Braun, B. R., Nguyen, L. H., and Geiduschek, E. P. (1990). S. cerevisiae TFIIIB is the transcription initiation factor proper of RNA polymerase III, while TFIIIA and TFIIIC are assembly factors. Cell 60, 235-245. |
| 945: | Kassavetis, G. A., Letts, G. A., and Geiduschek, E. P. (1999). A minimal RNA polymerase III transcription system.. EMBO J. 18, 5042-5051. |