7. Substitution of sigma factors may control initiation
9.7 Substitution of sigma factors may control initiation |
 |
Figure 9.18 E. coli sigma factors recognize promoters with different consensus sequences. (Numbers in the name of a factor indicate its mass.) |
The division of labors between a core enzyme that undertakes chain elongation and a sigma factor involved in site selection immediately raises the question of whether there is more than one type of sigma, each specific for a different class of promoters. Changes in sigma factors appear in some cases when there is a wholesale reorganization of transcription, for example, during the change in lifestyle that occurs in a sporulating bacterium (see next section). E. coli does not undergo such dramatic changes, but does use alternative sigma factors to respond to general environmental changes. They are listed in Figure 9.18. (They are named either by molecular weight of the product or for the gene.) The general factor, responsible for transcription of most genes under normal conditions, is σ70. The alternative sigma factors σ32, σE, and σ54 are activated in response to environmental changes; σ28 is used for expression of flagellar genes during normal growth, but its level of expression responds to changes in the environment. All the sigma factors except σ54 belong to the same protein family and function in the same general manner described earlier.
A common type of response to heat shock occurs in many organisms, both prokaryotic and eukaryotic. Upon an increase in temperature, synthesis of the proteins currently being made is turned off or down, and a new set of proteins is synthesized. The new proteins are the products of the heat shock genes. They play a role in protecting the cell against environmental stress, and are synthesized in response to other conditions as well as heat shock. Several of the heat shock proteins are chaperones. In E. coli, the expression of 17 heat shock proteins is triggered by changes at transcription. The gene rpoH is a regulator needed to switch on the heat shock response. Its product is σ32, which functions as an alternative sigma factor that causes transcription of the heat shock genes (499).
The heat shock response is accomplished by increasing the amount of σ32 when the temperature increases, and decreasing its activity when the temperature change is reversed. The basic signal that induces production of σ32 is the accumulation of unfolded (partially denatured) proteins that results from increase in temperature. The σ32 protein is unstable, which is important in allowing its quantity to be increased or decreased rapidly. σ70 and σ32 can compete for the available core enzyme, so that the set of genes transcribed during heat shock depends on the balance between them.
Another group of heat-regulated genes is controlled by the factor σE. It appears to respond to more extreme temperature shifts than σ32. It is induced by accumulation of unfolded proteins that are usually found in the periplasmic space or outer membrane. Less is known about this sigma factor and about the genes it controls.
Another sigma factor is used under conditions of nitrogen starvation. E. coli cells contain a small amount of σ54, which is activated when ammonia is absent from the medium. In these conditions, genes are turned on to allow utilization of alternative nitrogen sources. Counterparts to this sigma factor have been found in a wide range of bacteria, so it represents a response mechanism that has been conserved in evolution.
Another case of evolutionary conservation of sigma factors is presented by the factor σF, which is present in small amounts and causes RNA polymerase to transcribe genes involved in chemotaxis and flagellar structure. Its counterpart in B. subtilis is σD, which controls flagellar and motility genes; factors with the same promoter specificity are present in many species of bacteria.
Each sigma factor causes RNA polymerase to initiate at a particular set of promoters. By analyzing the sequences of these promoters, we can show that each set is identified by unique sequence elements. Indeed, the sequence of each type of promoter ensures that it is recognized only by RNA polymerase directed by the appropriate sigma factor. We can deduce the general rules for promoter recognition from the identification of the genes responding to the sigma factors found in E. coli and those involved in sporulation in B. subtilis (which are discussed in the next section).
A significant feature of the promoters for each enzyme is that they have the same size and location relative to the startpoint, and they show conserved sequences only around the usual centers of V35 and V10. The consensus sequences for each set of promoters are different from one another at either or both of the V35 and V0 positions. This means that an enzyme containing a particular sigma factor can recognize only its own set of promoters, so that transcription of the different groups is mutually exclusive. So substitution of one sigma factor by another turns off transcription of the old set of genes as well as turning on transcription of a new set of genes.
The definition of a series of different consensus sequences recognized at V35 and V10 by holoenzymes containing different sigma factors carries the immediate implication that the sigma subunit must itself contact DNA in these regions. This suggests the general principle that there is a common type of relationship between sigma and core enzyme, in which the sigma factor is positioned in such a way as to make critical contacts with the promoter sequences in the vicinity of V35 and V10.
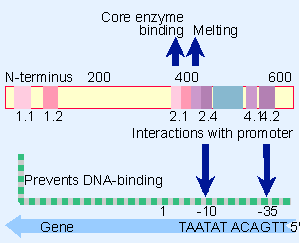 |
Figure 9.19 A map of the E. coli s70 factor identifies conserved regions. Regions 2.1 and 2.2 contact core polymerase, 2.3 is required for melting, and 2.4 and 4.2 contact the -10 and -35 promoter elements. The N-terminal region prevents 2.4 and 4.2 from binding to DNA in the absence of core enzyme. |
Comparisons of the sequences of several bacterial sigma factors identify regions that have been conserved. Their locations in E. coli σ70 are summarized in Figure 9.19 (503).
Direct evidence that sigma contacts the promoter directly at both the V35 and V10 consensus sequences is provided by mutations in sigma that suppress mutations in the consensus sequences. When a mutation at a particular position in the promoter prevents recognition by RNA polymerase, and a compensating mutation in sigma factor allows the polymerase to use the mutant promoter, we may conclude that the relevant base pair in DNA is contacted by the amino acid that has been substituted. Figure 9.19 shows that two short parts of regions 2 and 4 (named 2.4 and 4.2) are involved in contacting bases in the V10 and V35 elements, respectively. Both of these regions form short stretches of α-helix in the protein. Experiments with heteroduplexes show that σ70 makes contacts with bases principally on the nontemplate strand, and it continues to hold these contacts after the DNA has been unwound in this region. This suggests that sigma factor could be important in the melting reaction.
Region 2.3 resembles proteins that bind single-stranded nucleic acids, and is involved in the melting reaction. Regions 2.1 and 2.2 (which is the most highly conserved part of sigma) are involved in the interaction with core enzyme. It is assumed that all sigma factors bind the same regions of the core polymerase (ensuring that the reactions are competitive).
When the N-terminal region of σ70 is removed, the shortened protein becomes able to bind specifically to promoter sequences. This suggests that the N-terminal region behaves as an autoinhibition domain. It occludes the DNA-binding domains when σ70 is free. Association with core enzyme changes the conformation of sigma so that the inhibition is released, and the core domain becomes able to bind to DNA. When sigma binds to the core polymerase, the autoinhibition domain swings ~20 Å away from the DNA-binding domains, and the DNA-binding domains separate from one another by ~15 Å, presumably to acquire a more elongated conformation appropriate for contacting DNA. Mutations in either the V10 or V35 sequences prevent an (N-terminal-deleted) σ70 from binding to DNA, which suggests that σ70 contacts both sequences simultaneously. This implies that the sigma factor must have a rather elongated structure, extending over the ~68 Å of two turns of DNA.
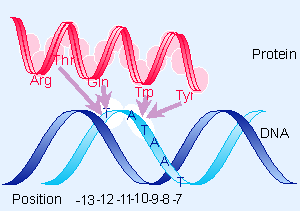 |
Figure 9.20 Amino acids in the 2.4 a-helix of s70 contact specific bases in the nontemplate strand of the -10 promoter sequence. |
The use of α-helical motifs in proteins to recognize duplex DNA sequences is common, as we see in more detail in Chapters 10 and 11. Amino acids separated by 3 V4 positions lie on the same face of an α-helix and are therefore in a position to contact adjacent base pairs. Figure 9.20 shows that amino acids lying along one face of the 2.4 region α-helix contact the bases at positions V12 to V10 of the V10 promoter sequence.
An interesting difference in behavior is found with the σ54 factor. This causes RNA polymerase to recognize promoters that have a distinct consensus sequence, with a conserved element at V10 and another close by at V20 (given in the " V35" column of Figure 9.18). So the geometry of the polymerase-promoter complex is different under the direction of this sigma factor. This is associated with a change in the pattern of regulation, in which sites that are rather distant from the promoter influence its activity, unlike the control of other promoters, where the regulator sites are always in close proximity to the promoter. The behavior of σ54 itself is different from other sigma factors, most notably in its ability to bind to DNA independently of core polymerase. In these regards, σ54 is more like the eukaryotic regulators we discuss in 20 Initiation of transcription than the typical prokaryotic regulators discussed in Chapters 10 and 11.
| Research | |
| 499: | Grossman, A. D., Erickson, J. W., and Gross, C. A. (1984). The htpR gene product of E. coli is a sigma factor for heat-shock promoters. Cell 38, 383-390. |
| 503: | Dombrowski, A. J. et al. (1992). Polypeptides containing highly conserved regions of transcription initiation factor s70 exhibit specificity of binding to promoter DNA. Cell 70, 501-512. |