5. Anterior development uses localized gene regulators
29.4 Maternal gene products establish gradients in early embryogenesis |
| Key terms defined in this section |
| Morphogen is a factor that induces development of particular cell types in a manner that depends on its concentration. |
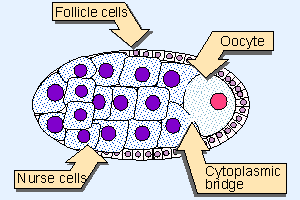 |
Figure 29.3 A Drosophila follicle contains an outer surface of follicle cells that surround nurse cells that are in close contact with the oocyte. Nurse cells are connected by cytoplasmic bridges to each other and to the anterior end of the oocyte. Follicle cells are somatic; nurse cells and the oocyte are germline in origin. |
An initial asymmetry is imposed on the Drosophila oocyte during oogenesis. Figure 29.3 illustrates the structure of a follicle in the Drosophila ovary. A single progenitor undergoes four successive mitoses to generate 16 interconnected cells. The connections are known as "cytoplasmic bridges" or "ring canals." Individual cells have 2, 3, or 4 such connections. One of the two cells that has 4 connections undergoes meiosis to become the oocyte; the other 15 cells become "nurse cells." Cytoplasmic material, including protein and RNA, passes from the nurse cells to the oocyte; the accumulation of such material accounts for a considerable part of the volume of the egg. The cytoplasmic connections are made at one end of the oocyte, and this end becomes the anterior end of the egg.
Genes that are expressed within the mother fly are important for early development. These maternal genes are identified by female sterile mutations. They do not affect the mother itself, but are required in order to have progeny. Females with such mutations lay eggs that fail to develop into adults; the embryos can be recognized by defects in the cuticular pattern, and they die during development.
The common feature in all maternal genes is that they are expressed prior to fertilization (although their products may act either at the time of expression or be stored for later use). The maternal genes are divided into two classes, depending on their site of expression. Genes that are expressed in somatic cells of the mother that affect egg development are called maternal somatic genes. For example, they may act in the follicle cells. Genes that are expressed within the germline are called maternal germline genes. These genes may act either in the nurse cell or the oocyte. Some genes act at both stages.
Four groups of genes concerned with the development of particular regions of the embryo can be identified by mutations in maternal genes. The genes in each group can be organized into a pathway that reflects their order of action, by conventional genetic tests (such as comparing the properties of double mutants with the individual mutants) or by biochemical assays (showing which mutants contain components that can bypass the stages that are blocked in other mutants; for review see Mahowald and Hardy, 1985; Lawrence, 1992).
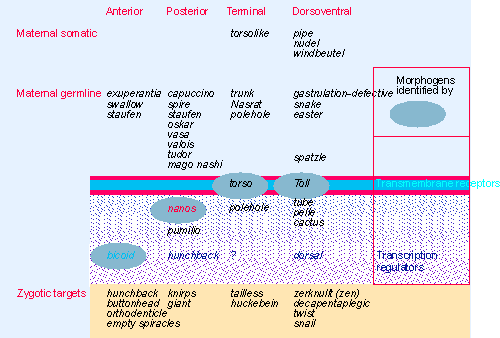 |
Figure 29.4 Each of the four maternal systems that functions in the egg is initiated outside the egg. The pathway is carried into the egg, where each pathway has a localized product that is the morphogen. This may be a receptor or a regulator of gene expression. The final component is a transcription factor, which acts on zygotic targets that are responsible for the next stage of development. |
The components of these pathways are summarized in Figure 29.4, which shows that there is a common principle to their operation. Each pathway is initiated by localized events outside the egg; this results in the localization of a signal within the egg. This signal takes the form of a protein with an asymmetric distribution; this is called a morphogen. Formally, we may define a morphogen as a protein whose local concentration (or activity) causes the surrounding region to take up a particular structure or fate. In each of these systems, the morphogen either is a transcriptional regulator or leads to the activation of a transcription factor in the localized region. Three systems are concerned with the anterior-posterior axis, and one with the dorsal-ventral axis:
- The anterior system is responsible for development of the head and thorax. The maternal germline products are required to localize the bicoid product at the anterior end of the egg. In fact, bicoid mRNA is transcribed in nurse cells and transported into the oocyte. Bicoid protein is the morphogen: it functions as a transcriptional regulator, and controls expression of the gene hunchback (and probably also other segmentation and homeotic genes).
- The posterior system is responsible for the segments of the abdomen. The nature of the initial asymmetrical event is not clear. A large number of products act to cause the localization of the product of nanos, which is the morphogen. This leads to localized repression of expression of hunchback (via control of translation of the mRNA).
- The terminal system is responsible for development of the specialized structures at the unsegmented ends of the egg (the acron at the head, and the telson at the tail). As indicated by the dependence on maternal somatic genes, the initial events that create asymmetry occur in the follicle cells. They lead to localized activation of the transmembrane receptor coded by torso; the end product of the pathway has yet to be identified.
- The fourth system is responsible for dorsal-ventral development. The pathway is initiated by a signal from a follicle cell on the ventral side of the egg. It is transmitted through the transmembrane receptor coded by Toll. This leads to a gradient of activation of the transcription factor produced by dorsal (by controlling its localization within the cell).
About 30 maternal genes involved in pattern formation have been identified. All of the components of the four pathways are maternal, so we see that the systems for establishing the initial pattern formation all depend on events that occur prior to fertilization. The two body axes are established independently. Mutations that affect polarity cause posterior regions to develop as anterior structures, or ventral regions to develop in dorsal form. On the anterior-posterior axis, the anterior and posterior systems provide opposing gradients, with sources at the anterior and posterior ends of the embryo, respectively, that control development of the segments of the body. Defects in either system affect the body segments. The terminal and dorsal-ventral systems operate independently of the other systems (for review see Ingham, 1988).
| Reviews | |
| Ingham, P. W. (1988). The molecular genetics of embryonic pattern formation in Drosophila. Nature 335, 25-34. | |
| Lawrence, P. (1992). The Making of a Fly. Blackwell Scientific, Oxford. | |
| Lawrence, P. (1992). The Making of a Fly. Blackwell Scientific, Oxford. | |
| Mahowald, A. P. and Hardy, P. A. (1985). Genetics of Drosophila embryogenesis. Ann. Rev. Genet. 19, 149-177. | |