11. Repair systems correct damage to DNA
14.11 Repair systems correct damage to DNA |
Injury to DNA is minimized by systems that recognize and correct the damage. The repair systems are likely to be as complex as the replication apparatus itself, which indicates their importance for the survival of the cell. The measured rate of mutation reflects a balance between the number of damaging events occurring in DNA and the number that have been corrected (or miscorrected).
"Damage" to DNA consists of any change introducing a deviation from the usual double-helical structure. We can divide such changes into two general classes:
- Single base changes affect the sequence but not the overall structure of DNA. They do not affect transcription or replication, when the strands of the DNA duplex are separated. So these changes exert their damaging effects on future generations through the consequences of the change in DNA sequence (see 1 Genes are DNA). Such an effect is caused by the conversion of one base into another that is not properly paired with the partner base. Figure 14.25 gives two examples: deamination of cytosine (spontaneously or by chemical mutagen) creates a mismatched U PG pair; while a replication error that inserts adenine instead of cytosine creates an A PG pair. Similar consequences could result from covalent addition of a small group to a base that modifies its ability to base pair. These changes may result in very minor structural distortion (as in the case of a U PG pair) or quite significant change (as in the case of an A PG pair), but the common feature is that the mismatch persists only until the next replication.
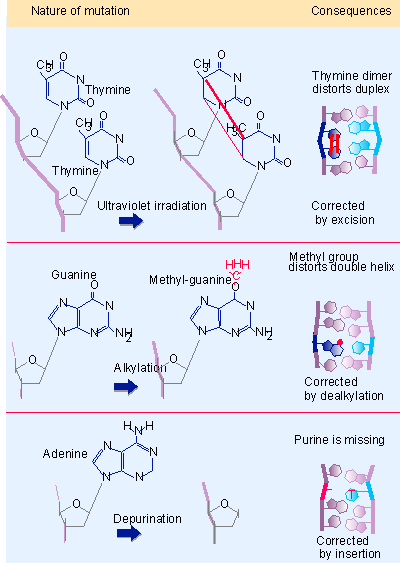
Figure 14.26 Modifications or removal of bases may cause structural defects that prevent replication or induce mutations in each replication cycle until they are removed. - Structural distortions may provide a physical impediment to replication or transcription. Figure 14.26 shows some examples. Introduction of covalent links between bases on one strand of DNA or between bases on opposite strands inhibits replication and transcription. A well studied example of a structural distortion is caused by ultraviolet irradiation, which introduces covalent bonds between two adjacent thymine bases, giving the intrastrand pyrimidine dimer drawn in the figure. Similar consequences could result from addition of a bulky adduct to a base that distorts the structure of the double helix. A single-strand nick or the removal of a base prevents a strand from serving as a proper template for synthesis of RNA or DNA. The common feature in all these changes is that the damaged adduct remains in the DNA, continuing to cause structural problems and/or induce mutations, until it is removed.
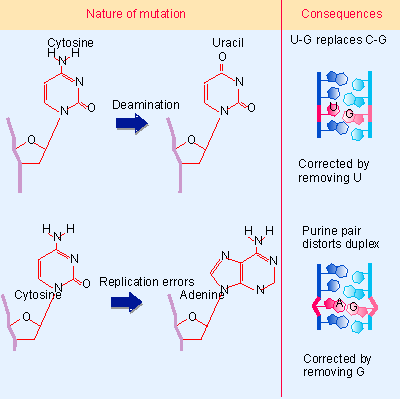 |
Figure 14.25 Substitutions of individual bases create mismatched pairs that may be corrected by replacing one base; if uncorrected they cause a mutation in one daughter duplex. |
Repair systems often can recognize a range of distortions in DNA as signals for action, and a cell may have several systems able to deal with DNA damage. We may divide them into several general types:
- Direct repair is rare and involves the reversal or simple removal of the damage. Photoreactivation of pyrimidine dimers, in which the offending covalent bonds are reversed by a light Vdependent enzyme, is the best example. This system is widespread in nature, and appears to be especially important in plants. In E. coli it depends on the product of a single gene (phr) that codes for an enzyme called photolyase.
- Excision repair is initiated by a recognition enzyme that sees an actual damaged base or a change in the spatial path of DNA. Recognition is followed by excision of a sequence that includes the damaged bases; then a new stretch of DNA is synthesized to replace the excised material. Such systems are common; some recognize general damage to DNA, while others act upon specific types of base damage (glycosylases remove specific altered bases; AP endonucleases remove residues from sites at which purine bases have been lost). There are often multiple excision repair systems in a single cell type, and they probably handle most of the damage that occurs.
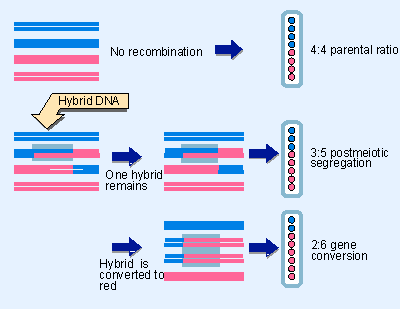
Figure 14.14 Spore formation in the Ascomycetes allows determination of the genetic constitution of each of the DNA strands involved in meiosis. - Mismatch repair is accomplished by scrutinizing DNA for apposed bases that do not pair properly. Mismatches that arise during replication are corrected by distinguishing between the "new" and "old" strands and preferentially correcting the sequence of the newly synthesized strand. Mismatches also occur when hybrid DNA is created during recombination, and their correction upsets the ratio of parental alleles (see Figure 14.14). Other systems deal with mismatches generated by base conversions, such as the result of deamination. The importance of these systems is emphasized by the fact that cancer is caused in human populations by mutation of genes related to those involved in mismatch repair in yeast.
- Tolerance systems cope with the difficulties that arise when normal replication is blocked at a damaged site. They provide a means for a damaged template sequence to be copied, probably with a relatively high frequency of errors. They are especially important in higher eukaryotic cells.
- Retrieval systems comprise another type of tolerance system. When damage remains in a daughter molecule, and replication has been forced to bypass the site, a retrieval system uses recombination to obtain another copy of the sequence from an undamaged source. These "recombination-repair" systems are well characterized in bacteria; it is not clear how important they are elsewhere.
Mutations that affect the ability of E. coli cells to engage in DNA repair fall into groups, which correspond to several repair pathways (not necessarily all independent). The major known pathways are the uvr excision repair system, the dam replication mismatch-repair system, and the recB and recF recombination and recombination-repair pathways.
When the repair systems are eliminated, cells become exceedingly sensitive to ultraviolet irradiation. The introduction of UV-induced damage has been a major test for repair systems, and so in assessing their activities and relative efficiencies, we should remember that the emphasis might be different if another damaged adduct were studied.
- Chapter IV How Consumers Think About Interactive Aspects of Web Advertising
- Chapter IX Extrinsic Plus Intrinsic Human Factors Influencing the Web Usage
- Chapter XI User Satisfaction with Web Portals: An Empirical Study
- Chapter XV Customer Trust in Online Commerce
- Chapter XVIII Web Systems Design, Litigation, and Online Consumer Behavior