6. Posterior development uses another localized regulator
29.5 Anterior development uses localized gene regulators |
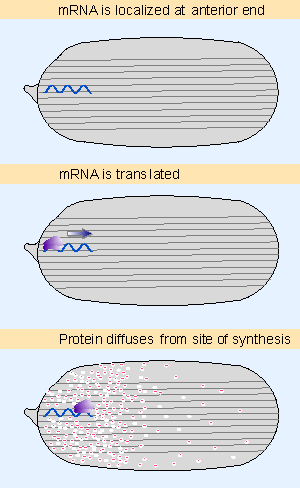 |
Figure 29.5 Translation of a localized mRNA generates a gradient of protein as the products diffuses away from the site of synthesis. Animated figure |
Establishing asymmetry in an egg requires that some components Xeither RNAs or proteins Xare localized instead of being diffused evenly through the cytosol. In anterior-posterior development in Drosophila, certain mRNAs are localized at the anterior or posterior end. Figure 29.5 shows that when they are translated, their protein products diffuse away from the ends of the egg, generating a gradient along the anterior-posterior axis.
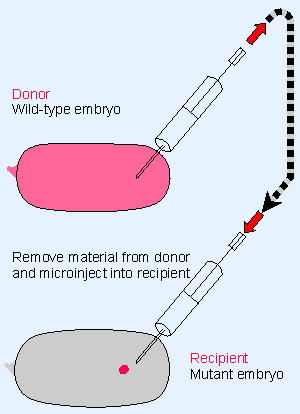 |
Figure 29.6 Mutant embryos that cannot develop can be rescued by injecting cytoplasm taken from a wild-type embryo. The donor can be tested for time of appearance and location of the rescuing activity; the recipient can be tested for time at which it is susceptible to rescue and the effects of injecting material at different locations. |
The existence of localized concentrations of materials needed for development can be tested by the rescue protocol summarized in Figure 29.6. Material is removed from a wild-type embryo and injected into the embryo of a mutant that is defective in early development. If the mutant embryo develops normally, we may conclude that the mutation causes a deficiency of material that is present in the wild-type embryo. This allows us to distinguish components that are necessary for morphogenesis, or that are upstream in the pathway, from the morphogen itself Xonly the morphogen has the property of localized rescue.
The rescue technique identifies bicoid as the morphogen required for anterior development. bicoid mutants do not develop heads; but the defect can be remedied by injecting mutant eggs with cytoplasm taken from the anterior tip of a wild-type embryo. Indeed, anterior structures develop elsewhere in the mutant embryo if wild-type anterior cytoplasm is injected! (This is called ectopic expression.) The extent of the rescue depends on the amount of wild-type cytoplasm injected. And the efficacy of the donor cytoplasm depends on the number of wild-type bicoid genes carried by the donor. This suggests that the anterior region of a wild-type embryo contains a concentration of some product that depends on the bicoid gene dosage. By purifying the active component in the preparation, it is possible to show that purified bicoid mRNA can substitute for the anterior cytoplasm. These results imply that the components on which bicoid acts are ubiquitous, and all that is required to trigger formation of anterior structures is an appropriate concentration of bicoid product.
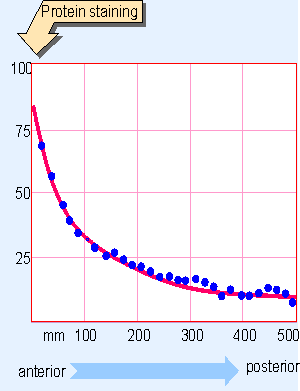 |
Figure 29.7 Bicoid protein forms a gradient during D. melanogaster development that extends for ~200 mm along the egg of 500 mm. |
The product of bicoid establishes a gradient with its source (and therefore the highest concentration) at the anterior end of the embryo. The RNA is localized at the anterior tip of the embryo, but it is not translated during oogenesis. Translation begins soon after fertilization. The protein then establishes a gradient along the embryo, as indicated in Figure 29.7. The gradient could be produced by diffusion of the protein product from the localized source at the anterior tip. The gradient is established by division 7, and remains stable until after the blastoderm stage.
What is the consequence of establishing the bicoid gradient? The gradient can be increased or decreased by changing the number of functional gene copies in the mother. The concentration of bicoid protein is correlated with the development of anterior structures. Weakening the gradient causes anterior segments to develop more posterior-like characteristics; strengthening the gradient causes anterior-like structures to extend farther along the embryo. So the bicoid protein behaves as a morphogen that determines anterior-posterior position in the embryo in a concentration-dependent manner.
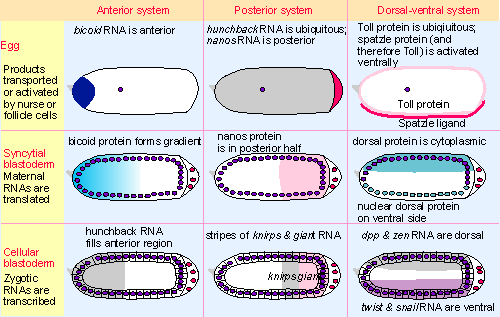 |
Figure 29.17 In each axis-determining system, localized products in the egg cause other maternal RNAs or proteins to be broadly localized at syncytial blastoderm, and zygotic RNAs are transcribed in bands at cellular blastoderm. |
The fate of cells in the anterior part of the embryo is determined by the concentration of bicoid protein in which they find themselves. The bicoid product is a sequence-specific DNA-binding protein that regulates transcription of genes to whose promoters it binds. The immediate effect of bicoid is exercised on other genes that in turn regulate the development of yet further genes. A major target for bicoid is the gene hunchback (see later). Transcription of hunchback is turned on by bicoid in a dose-dependent manner, that is, hunchback is activated above a certain threshold of bicoid protein. This allows a gradient to provide a spatial on-off switch that affects gene expression. In this way, quantitative differences in the amount of the morphogen (bicoid protein) are transformed into qualitatively different states (cell structures) during embryonic development. bicoid plays an instructive role in anterior development, since it is a positive regulator that is needed for expression of genes that in turn determine the synthesis of anterior structures. The effect of bicoid on hunchback is to produce a band of expression that occupies the anterior part of the embryo (see Figure 29.17; for review see McGinnis and Krumlauf, 1992).
| Reviews | |
| McGinnis, W. and Krumlauf, R. (1992). Homeobox genes and axial patterning. Cell 68, 283-302. | |