2. Clonal selection amplifies lymphocytes that respond to individual antigens
24.1 Introduction |
| Key terms defined in this section |
| Antigen is any molecule whose entry into an organism provokes synthesis of an antibody (immunoglobulin). Superfamily is a set of genes all related by presumed descent from a common ancestor, but now showing considerable variation. T cells are lymphocytes of the T (thymic) lineage; may be subdivided into several functional types. They carry TcR (T-cell receptor) and are involved in the cell-mediated immune response. |
It is an axiom of genetics that the genetic constitution created in the zygote by the combination of sperm and egg is inherited by all somatic cells of the organism. We look to differential control of gene expression, rather than to changes in DNA content, to explain the different phenotypes of particular somatic cells.
Yet there are exceptional situations in which the reorganization of certain DNA sequences is used to regulate gene expression or to create new genes. The immune system provides a striking and extensive case in which the content of the genome changes, when recombination creates active genes in lymphocytes. Other cases are represented by the substitution of one sequence for another to change the mating type of yeast or to generate new surface antigens by trypanosomes (see 17 Rearrangement of DNA).
The immune response of vertebrates provides a protective system that distinguishes foreign proteins from the proteins of the organism itself. Foreign material (or part of the foreign material) is recognized as comprising an antigen. Usually the antigen is a protein (or protein-attached moiety) that has entered the bloodstream of the animal Xfor example, the coat protein of an infecting virus. Exposure to an antigen initiates production of an immune response that specifically recognizes the antigen and destroys it.
Immune reactions are the responsibility of white blood cells Xthe B and T lymphocytes, and macrophages. The lymphocytes are named after the tissues that produce them. In mammals, B cells mature in the bone marrow, while T cells mature in the thymus. Each class of lymphocyte uses the rearrangement of DNA as a mechanism for producing the proteins that enable it to participate in the immune response.
The immune system has many ways to destroy an antigenic invader, but it is useful to consider them in two general classes. Which type of response the immune system mounts when it encounters a foreign structure depends partly on the nature of the antigen. The response is defined according to whether it is executed principally by B cells or T cells.
The humoral response depends on B cells. It is mediated by the secretion of antibodies, which are immunoglobulin proteins. Production of an antibody specific for a foreign molecule is the primary event responsible for recognition of an antigen. Recognition requires the antibody to bind to a small region or structure on the antigen.
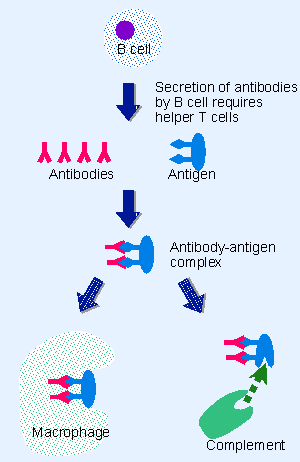 |
Figure 24.1 Humoral immunity is conferred by the binding of free antibodies to antigens to form antigen-antibody complexes that are removed from the bloodstream by macrophages or that are attacked directly by the complement proteins. |
The function of antibodies is represented in Figure 24.1. Foreign material circulating in the bloodstream, for example, a toxin or pathogenic bacterium, has a surface that presents antigens. The antigen(s) are recognized by the antibodies, which form an antigen-antibody complex. This complex then attracts the attention of other components of the immune system.
The humoral response depends on these other components in two ways. First, B cells need signals provided by T cells to enable them to secrete antibodies. These T cells are called helper T cells, because they assist the B cells. Second, antigen-antibody formation is a trigger for the antigen to be destroyed. The major pathway is provided by the action of complement, a component whose name reflects its ability to "complement" the action of the antibody itself. Complement consists of a set of ~20 proteins that function through a cascade of proteolytic actions. If the target antigen is part of a cell, for example, an infecting bacterium, the action of complement culminates in lysing the target cell. The action of complement also provides a means of attracting macrophages, which scavenge the target cells or their products. Alternatively, the antigen-antibody complex may be taken up directly by macrophages (scavenger cells) and destroyed.
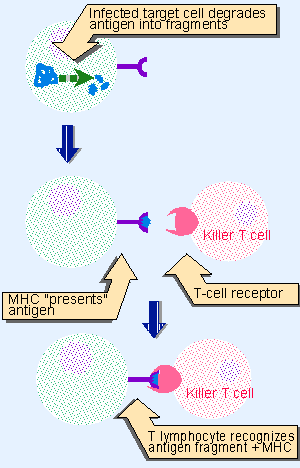 |
Figure 24.2 In cell-mediated immunity, killer T cells use the T-cell receptor to recognize a fragment of the foreign antigen which is presented on the surface of the target cell by the MHC protein. |
The cell-mediated response is executed by a class of T lymphocytes called cytotoxic T cells (also called killer T cells). The basic function of the T cell in recognizing a target antigen is indicated in Figure 24.2. A cell-mediated response typically is elicited by an intracellular parasite, such as a virus that infects the body’s own cells. As a result of the viral infection, foreign (viral) antigens are displayed on the surface of the cell. These antigens are recognized by the T-cell receptor (TCR), which is the T cells’ equivalent of the antibody produced by a B cell.
A crucial feature of this recognition reaction is that the antigen must be presented by a cellular protein that is a member of the MHC (major histocompatibility complex). The MHC protein has a groove on its surface that binds a peptide fragment derived from the foreign antigen. The combination of peptide fragment and MHC protein is recognized by the T-cell receptor. Every individual has a characteristic set of MHC proteins. They are important in graft reactions; a graft of tissue from one individual to another is rejected because of the difference in MHC proteins between the donor and recipient, an issue of major medical importance. The demand that the T lymphocytes recognize both foreign antigen and MHC protein ensures that the cell-mediated response acts only on host cells that have been infected with a foreign antigen. (We discuss the division of MHC proteins into the general types of class I and class II later in this chapter.)
The purpose of each type of immune response is to attack a foreign target. Target recognition is the prerogative of B-cell immunoglobulins and T-cell receptors. A crucial aspect of their function lies in the ability to distinguish "self" from "nonself." Proteins and cells of the body itself must never be attacked. Foreign targets must be destroyed entirely. The property of failing to attack "self" is called tolerance. Loss of this ability results in an autoimmune disease, in which the immune system attacks its own body, often with disastrous consequences.
What prevents the lymphocyte pool from responding to "self" proteins? Tolerance probably arises early in lymphocyte cell development when B and T cells that recognize "self" antigens are destroyed. This is called clonal deletion. In addition to this negative selection, there is also positive selection for T cells carrying certain sets of T-cell receptors.
A corollary of tolerance is that it can be difficult to obtain antibodies against proteins that are closely related to those of the organism itself. As a practical matter, therefore, it may be difficult to use (for example) mice or rabbits to obtain antibodies against human proteins that have been highly conserved in mammalian evolution. The tolerance of the mouse or rabbit for its own protein may extend to the human protein in such cases.
Each of the three groups of proteins required for the immune response Ximmunoglobulins, T-cell receptors, MHC proteins Xis diverse. Examining a large number of individuals, we find many variants of each protein. Each protein is coded by a large family of genes; and in the case of antibodies and the T-cell receptors, the diversity of the population is increased by DNA rearrangements that occur in the relevant lymphocytes.
Immunoglobulins and T-cell receptors are direct counterparts, each produced by its own type of lymphocyte. The proteins are related in structure, and their genes are related in organization. The sources of variability are similar. The MHC proteins also share some common features with the antibodies, as do other lymphocyte-specific proteins. In dealing with the genetic organization of the immune system, we are therefore concerned with a series of related gene families, indeed a superfamily that may have evolved from some common ancestor representing a primitive immune response.