2. Response elements identify genes under common regulation
21.2 Response elements identify genes under common regulation |
The principle that emerges from characterizing groups of genes under common control is that they share a promoter (or enhancer) element that is recognized by a regulatory transcription factor. An element that causes a gene to respond to such a factor is called a response element; examples are the HSE (heat shock response element), GRE (glucocorticoid response element), SRE (serum response element).
Response elements have the same general characteristics as upstream elements of promoters or enhancers. They contain short consensus sequences, and copies of the response elements found in different genes are closely related, but not necessarily identical. The region bound by the factor extends for a short distance on either side of the consensus sequence. In promoters, the elements are not present at fixed distances from the startpoint, but are usually <200 bp upstream of it. The presence of a single element usually is sufficient to confer the regulatory response, but sometimes there are multiple copies.
Response elements may be located in promoters or in enhancers. Some types of elements are typically found in one rather than the other: usually an HSE is found in a promoter, while a GRE is found in an enhancer. We assume that all response elements function by the same general principle. A gene is regulated by a sequence at the promoter or enhancer that is recognized by a specific protein. The protein functions as a transcription factor needed for RNA polymerase to initiate. Active protein is available only under conditions when the gene is to be expressed.
An example of a situation in which many genes are controlled by a single factor is provided by the heat shock response. This is common to a wide range of prokaryotes and eukaryotes and involves multiple controls of gene expression: an increase in temperature turns off transcription of some genes, turns on transcription of the heat shock genes, and causes changes in the translation of mRNAs. The control of the heat shock genes illustrates the differences between prokaryotic and eukaryotic modes of control. In bacteria, a new sigma factor is synthesized that directs RNA polymerase holoenzyme to recognize an alternative V10 sequence common to the promoters of heat shock genes (see 9 Transcription). In eukaryotes, the heat shock genes also possess a common consensus sequence (HSE), but it is located at various positions relative to the startpoint, and is recognized by an independent transcription factor, HSTF. The activation of this factor therefore provides a means to initiate transcription at the specific group of ~20 genes that contains the appropriate target sequence at its promoter.
All the heat shock genes of D. melanogaster contain multiple copies of the HSE. The HSTF binds cooperatively to adjacent response elements. Both the HSE and HSTF have been conserved in evolution, and it is striking that a heat shock gene from D. melanogaster can be activated in species as distant as mammals or sea urchins. The HSTF proteins of fruit fly and yeast appear similar, and show the same footprint pattern on DNA containing HSE sequences. Yeast HSTF becomes phosphorylated when cells are heat-shocked; this modification is responsible for activating the protein.
The metallothionein (MT) gene provides an example of how a single gene may be regulated by many different circuits. The metallothionein protein protects the cell against excess concentrations of heavy metals, by binding the metal and removing it from the cell. The gene is expressed at a basal level, but is induced to greater levels of expression by heavy metal ions (such as cadmium) or by glucocorticoids. The control region combines several different kinds of regulatory elements, and suggests the principle that when a promoter is regulated in more than one way, each regulatory event depends on binding of its own protein to a particular sequence.
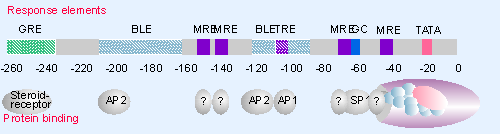 |
Figure 21.1 The regulatory region of a human metallothionein gene contains regulator elements in both its promoter and enhancer. The promoter has elements for metal induction; an enhancer has an element for response to glucocorticoid. Promoter elements are shown above the map, and proteins that bind them are indicated below. |
The organization of the promoter for a MT gene is summarized in Figure 21.1. A major feature of this map is the high density of elements that can activate transcription. The TATA and GC boxes are located at their usual positions fairly close to the startpoint. Also needed for the basal level of expression are the two basal level elements (BLE), which fit the formal description of enhancers. Although located near the startpoint, they can be moved elsewhere without loss of effect. They contain sequences related to those found in other enhancers, and are bound by proteins that bind the SV40 enhancer.
The TRE is a consensus sequence that is present in several enhancers, including one BLE of metallothionein and the 72 bp repeats of the virus SV40. The TRE has a binding site for factor AP1; this interaction is part of the mechanism for constitutive expression, in line with our previous description of AP1 as an upstream factor. However, AP1 binding also has a second function. The TRE confers a response to phorbol esters such as TPA (an agent that promotes tumors), and this response is mediated by the interaction of AP1 with the TRE. This binding reaction is one (not necessarily the sole) means by which phorbol esters trigger a series of transcriptional changes.
The inductive response to metals is conferred by the multiple MRE sequences, which function as promoter elements. The presence of one MRE confers the ability to respond to heavy metal; a greater level of induction is achieved by the inclusion of multiple elements.
The response to steroid hormones is governed by a GRE, located 250 bp upstream of the startpoint, which behaves as an enhancer. Deletion of this region does not affect the basal level of expression or the level induced by metal ions. But it is absolutely needed for the response to steroids.
The regulation of metallothionein illustrates the general principle that any one of several different elements, located in either an enhancer or promoter, can independently activate the gene. The absence of an element needed for one mode of activation does not affect activation in other modes. The variety of elements, their independence of action, and the apparently unlimited flexibility of their relative arrangements, suggest that a factor binding to any one element is able independently to increase the efficiency of initiation by the basal transcription apparatus, probably by virtue of protein-protein interactions that stabilize or otherwise assist formation of the initiation complex.