15. Transport receptors carry cargo proteins through the pore
8.15 Transport receptors carry cargo proteins through the pore |
| Key terms defined in this section |
| Exportins are transport receptors that bind cargo in the nucleus, and translocate into the cytoplasm where they release the cargo. Importins are transport receptors that bind cargo in the cytoplasm, and translocate into the nucleus where they release the cargo. Nucleoporin was originally defined to describe the components of the nuclear pore complex that bind to the inhibitory lectins, but now is used to mean any component of the basic nuclear pore complex. Translocation of a chromosome describes a rearrangement in which part of a chromosome is detached by breakage and then becomes attached to some other chromosome. |
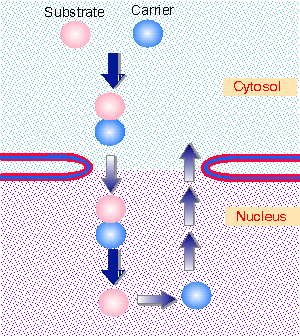 |
Figure 8.41 A carrier protein binds to a substrate, moves with it through the nuclear pore, is released on the other side, and must be returned for reuse. Animated figure |
The basic principle of import and export is illustrated in Figure 8.41. A carrier protein (or transport receptor) takes the substrate through the pore. The transport receptor must then be returned across the membrane to function in another cycle. The transport receptors are classified according to the direction in which they transport the cargo. Importins bind the cargo in the cytoplasm and release it in the nucleus. Exportins bind the cargo in the nucleus and release it in the cytoplasm.
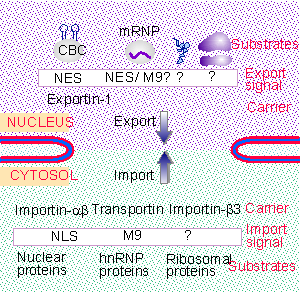 |
Figure 8.42 There are multiple pathways for nuclear export and import. |
There are multiple pathways for import and export. Transport for all of the substrates for any particular pathway can be inhibited by saturating that pathway with one of its substrates. Figure 8.42 summarizes the independent pathways. At least two different types of pathways exist for import of proteins; and each class of RNA is exported by a different system. Each transport receptor recognizes a particular type of sequence in its substrate, thus defining the specificity of the system (for review see Gorlich and Kutay, 1999).
 |
Figure 8.43 The assay for nuclear pore function uses permeabilized cells. |
The first handle on the process of import was provided by systems for transport that depend upon the presence of an NLS in the substrate protein. The complexity of the system makes it impossible to develop a true in vitro assay for nuclear pore import, but an effective system has been developed by using permeabilized cells. When cells are treated with digitonin, the plasma membrane becomes permeable, but the nuclear envelope remains intact. Labeled proteins can be imported into the nucleus in a process that is dependent upon the provision of cytosolic components (Adam, Marr, and Gerace, 1990). Figure 8.43 shows how this system has been used to characterize the transport process.
Transport can be divided into two stages: docking, which consists of binding to the pore; and translocation, which consists of movement through it. In the absence of ATP, proteins containing a nuclear import signal can bind at the pore, but they remain at the cytoplasmic face. A cytosolic fraction is needed for binding. When ATP is provided, proteins can be translocated through the pore. A different cytosolic fraction is needed to support translocation. The need for cytosolic fractions at both stages reinforces the view of the pore as a structure that provides the framework for transport, but that does not provide all of the necessary facilities for handling the substrates (Newmeyer and Forbes, 1988; Richardson et al., 1988; Moore and Blobel, 1992).
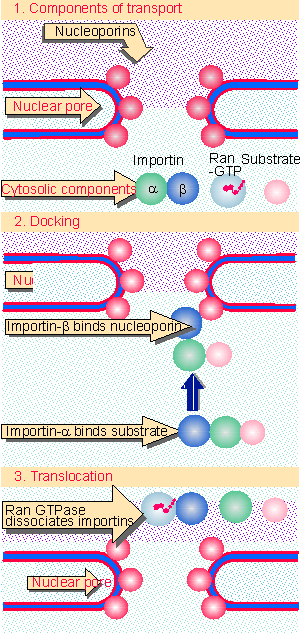 |
Figure 8.44 Nuclear import takes place in two stages. Both docking and translocation depend on cytosolic components. Translocation requires nucleoporins. |
Figure 8.44 summarizes the functions of the components involved in nuclear import. The transport receptor is the key intermediate in the docking reaction. It has the abilities to bind to the nuclear pore and to the cargo protein. In the best characterized example of an importin, the transport receptor is a dimer in which one subunit binds to the pore, and the other is an adaptor that binds to the cargo protein. The β subunit binds to the nuclear pore. The α subunit binds proteins that have an NLS sequence. The simplest import systems use transport receptors that are single subunits, such as transportin or importin-β3. In these cases, a single protein binds both to the cargo and to the nuclear pore (Moroianu et al., 1995). These transport receptors are related to importin-β, but they do not need an adaptor like importin-α to bind the substrate, that is, they are capable of binding the substrate directly.
Translocation through the pore is inhibited by wheat germ agglutinin, a lectin (glycoprotein). The component proteins of the pore that bind to lectins were originally called nucleoporins. Note that nucleoporin has since come to be used to mean any component of the nuclear pore complex. The lectin-binding components are localized at or near the region of the central pore, and appear to be located on both sides of the nuclear envelope. When they are removed, pore complexes remain normal in appearance, but can no longer function to transport large material. Material smaller than the pore size continues to be able to move through by diffusion. When the lectin-binding proteins are added back, they restore full activity to the deficient pores. This suggests that they are needed for active transport of material larger than the resting diameter. Some of these proteins have some simple peptide repeating motifs (GKFG, FG, FXFG), and it is probably these motifs that bind the importin-β carrier proteins (Radu et al., 1995). They are called FG-nucleoporins.
How does material move through the pore? One of the striking features of the composition of the nuclear pore complex is the lack of any protein with motor activity (Rout el al., 2000). This suggests that movement must depend on the affinity of the carrier proteins for the pore itself. But now we have to explain how transport can work for both import and export. The answer may lie in the properties of the monomeric G-protein, Ran.
The cytosolic fraction that supports translocation has two active components. One is Ran; the other may be involved in targeting Ran to the nuclear pore. Ran is a typical monomeric G-protein that can exist in either the GTP-bound or GDP-bound state (for introduction see G proteins). Its GTPase activity generates Ran-GDP. Then an exchange factor is needed to replace GDP with GTP to regenerate Ran-GTP.
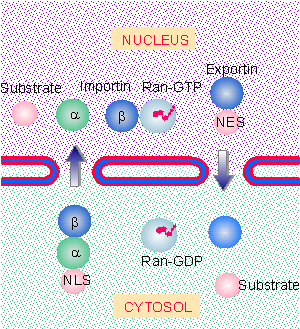 |
Figure 8.45 The state of the guanine nucleotide bound to Ran controls directionality of nuclear import and export. |
The directionality of nuclear import is controlled by the state of Ran (for review see Gorlich and Kutay, 1999). Figure 8.45 shows that conditions in the nucleus and cytosol differ so that typically there is Ran-GTP in the nucleus, but there is Ran-GDP in the cytosol. Export complexes are stable in the presence of Ran-GTP, whereas import complexes are stable in the presence of Ran-GDP. So export complexes are driven to form in the nucleus and dissociate in the cytosol, whereas the reverse is true of import complexes. The reaction has been best characterized for the complex of importin-αβ with an NLS-containing protein. The triple complex is stable in the presence of Ran-GDP, and thus can form in the cytosol. However, Ran-GTP causes importin-α to dissociate from importin-β. This leads to release of the substrate protein in the environment of the nucleus (Gorlich et al., 1996; Rexach and Blobel, 1995).
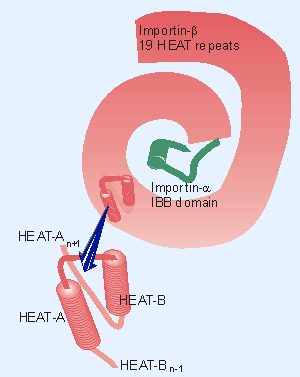 |
Figure 8.46 Importin-b consists of 19 HEAT repeats organized in a right-handed superhelix. Each HEAT unit consists of two a-helices (A and B) lying at an angle to one another. Importin-b is folded tightly around the IBB domain of importin-a. |
The crystal structures of complexes containing importin-β show that binding of importin-α is mutually exclusive with binding of Ran-GDP. Importin-β consists of a series of repeating units coiled into a superhelix. The individual repeating unit (called HEAT) itself consists of two α-helices (HEAT-A and HEAT-B). Importin-α has a similar structure, but in addition has a domain (IBB for importin-binding domain) that binds importin-β. Figure 8.46 shows that importin-β winds around the IBB of importin-α, making a tightly integrated dimer (Cingolani et al., 1999).
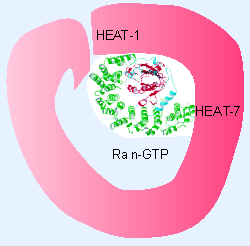 |
Figure 8.47 Importin-b binds Ran-GDP through close contacts to the N-terminal HEAT repeats and to repeats 7-8. The structure is significantly different from the importin-b importin-a structure. |
Figure 8.47 shows that Ran-GTP binds tightly to two of the HEAT repeats in importin-β. Its binding site partially overlaps the binding site of importin-α (Chook and Blobel, 1999). This explains why binding of Ran-GTP displaces importin-α (and the NLS of the cargo protein) from importin-β. There is a large structural change in Ran when GTP is hydrolyzed to GDP (involving the unfolding of a helical region), which explains why thedisplacement reaction is specific for Ran-GTP.
How does the importin-substrate complex cross the nuclear pore? FG-nucleoporins are distributed throughout the central pore. We do not know whether an importin-substrate complex moves across the pore continuously, or whether there is a series of docking and release events in which importin-substrate would bind to an FG-nucleoporin, dissociate, reassociate, bind to another FG-nucleoporin farther along the pore, and repeat the cycle. Position in the pore could be defined by the Ran-GTP/Ran-GDP gradient, but translocation as such does not appear to require GTP hydrolysis, and the source of energy for this movement is not known.
Export of mRNA has similar requirements to import. Mutations in a yeast FG-nucleoporin block export of RNA from the nucleus without affecting import of proteins. This suggests the possibility that the apparatus is similar for both import and export, but could have components that confer directionality or specificity for particular substrates. There is evidence for diversity in the export apparatus; using an assay for export of microinjected RNAs from the nucleus of the Xenopus oocyte, tRNAs, other small RNAs, and mRNAs appear to saturate transport only of their own class. This suggests that there are at least three groups of exported RNAs.
 |
Figure 8.48 The common feature in proteins that are exported from the nucleus to the cytosol is the presence of an NES. Multiple figure |
Export systems have similar components to import systems. Figure 8.48 illustrates some examples of export systems. The major substrates for export from the nucleus are ribonucleoproteins Xribosomes, mRNPs, snRNPs, and tRNA-protein complexes. In the first three cases, one of the protein components of the complex may be responsible for export (for example, for snRNAs it is the cap-binding complex (CBC). tRNA is bound directly by a specific export protein (Stade et al., 1997; Fornerod et al., 1997).
The NES binds to exportin-1, which is related in sequence to importin-β. Exportin-1 is needed for the export of U snRNAs, some proteins, and possibly (some) mRNAs. It binds the NES motif; it binds nucleoporins; and it binds Ran-GTP. The complex forms in the presence of Ran-GTP; hydrolysis of the GTP to generate Ran-GDP is accompanied by dissociation of the complex. So Ran controls directionality of export in the reverse sense from its control of import: because Ran-GTP is high in the nucleus, the complex forms there; because it becomes Ran-GDP in the cytosol, the complex dissociates there.
Some proteins are exported from the nucleus in particular circumstances; for example, the catalytic subunit of the enzyme cAPK (cyclic AMP-dependent kinase) functions in the nucleus. However, when an inhibitory subunit (PKI) is activated, it binds to the catalytic subunit, and the dimer is then exported from the nucleus. Some proteins are required to bind to RNAs and to export them from the nucleus; an example is the Rev protein of HIV, which causes export of certain RNAs from the nucleus.
As pointed out in Figure 8.41, in order to function more than once, an importin must return to the cytosol after taking its substrate into the nucleus. In effect, when an importin is released in the nucleus, it must become a substrate for an exportin! Such a reaction has been characterized for importin-α, which is bound in the presence of Ran-GTP by a protein called CAS (Kutay et al., 1997). CAS behaves in a similar manner to importin-β, except that it moves in the opposite direction. Like exportins, it dissociates from its substrate (importin-α) when Ran-GTP is hydrolyzed in the cytosol.
Some proteins "shuttle" between the cytoplasm and nucleus; they remain only briefly in either compartment before cycling back to the other. This behavior is characteristic of some proteins that are bound to poly(A)+ RNA in both the nucleus and the cytoplasm. The motif responsible for transport in one such protein (M9) has a single amino acid stretch that functions as both an import and export signal, and is therefore responsible for movement in both directions (for review see Gorlich and Mattaj, 1996).
5-22-2000
| Reviews | |
| Gorlich, D. and Kutay, U. (1999). Transport between the cell nucleus and cytoplasm.. Ann. Rev. Cell Dev. Biol. 15, 607-660. | |
| Gorlich, D. and Mattaj, I. W. (1996). Nucleocytoplasmic transport. Science 271, 1513-1518. | |
| Research | |
| Adam, S. A. , Marr, R. S. , and Gerace, L. (1990). Nuclear protein import in permeabilized mammalian cells requires soluble cytoplasmic factors.. J. Cell Biol. 111, 807-816. | |
| Chook, Y. M. and Blobel, G. (1999). Structure of the nuclear transport complex karyopherin-beta2-Ran x GppNHp. Nature 399, 230-237. | |
| Cingolani, G. , Petosa, C. , Weis, K. , Muller, C. W. , Cingolani, G. , Petosa, C. , Weis, K. , and Muller, C. W. (1999). Structure of importin-beta bound to the IBB domain of importin-alpha. Nature 399, 221-229. | |
| Fornerod, M. et al. (1997). CRM1 is an export receptor for leucine-rich nuclear export signals. Cell 90, 1051-1060. | |
| Gorlich, D. , Pante, N. , Kutay, U. , Aebi, U. , and Bischoff, F. R. (1996). Identification of different roles for RanGDP and RanGTP in nuclear protein import.. EMBO J. 15, 5584-5594. | |
| Kutay, U. et al. (1997). Export of importin a from the nucleus is mediated by a specific nuclear transport factor. Cell 90, 1061-1071. | |
| Moore, M. S. and Blobel, G. (1992). The two steps of nuclear import, targeting to the nuclear envelope and translocation through the nuclear pore, require different cytosolic factors. Cell 69, 939-950. | |
| Moroianu, J., Blobel, G., and Radu, A. (1995). Previously identified protein of uncertain function is importin a docks import substrate at nuclear pore complexes. Proc. Nat. Acad. Sci. USA 92, 2008-2011. | |
| Newmeyer, D. D. and Forbes, D. J. (1988). An NEM-sensitive cytosolic factor necessary for nuclear protein import: requirement in a signal-mediated binding to the nuclear pore. Cell 52, 641-653. | |
| Radu, A., Moore, M. S., and Blobel, G. (1995). The peptide repeat domain of nucleoporin Nup98 functions as a docking site in transport across the nuclear pore complex. Cell 81, 215-222. | |
| Rexach, M. and Blobel, G. (1995). Protein import into nuclei: association and dissociation reactions involving transport substrate, transport factors, and nucleoporins.. Cell 83, 683-692. | |
| Richardson, W. D. et al. (1988). Nuclear protein migration involves two steps: rapid binding at the nuclear envelope followed by slower translocation through nuclear pores. Cell 52, 655-664. | |
| Rout, M. P. et al. (2000). The yeast nuclear pore complex: composition, architecture, and transport mechanism.. J. Cell Biol. 148, 635-651. | |
| Stade, K. et al. (1997). Exportin 1 Crm1p is an essential nuclear export factor. Cell 90, 1041-1050. | |